Губы тритона и крылья цыпленка
На протяжении всего прошлого столетия биологи с помощью скальпелей, игл, пинцетов и всевозможных других инструментов резали, перетягивали, жгли, измельчали, скручивали и прокалывали эмбрионы, пытаясь обнаружить основные правила, по которым строятся животные. Пионеры эмбриологии целиком и полностью полагались на физические методы, когда перемещали и удаляли клетки, а затем анализировали аномалии формирования эмбриона. В результате этих "жестоких пыток" было получено несколько монстров, чьи поразительные признаки позволили выявить несколько ключевых закономерностей, регулирующих процессы развития животных[3].
В первую очередь среди всех ученых-эмбриологов того времени следует назвать Ганса Шпемана, который был первым и на протяжении шестидесяти с лишним лет единственным эмбриологом, удостоенным Нобелевской премии (впрочем, впоследствии, как мы с вами убедимся, эмбриологи наверстали упущенное). В одном из своих первых экспериментов Шпеман попытался установить, одинаковыми ли свойствами обладают две первые клетки эмбриона тритона. Шпеман взял тонкий волосок с головы своей дочери и туго затянул сделанную из него петлю между двумя первыми клетками, разделив эмбрион на две половинки. Каждая из клеток дала начало нормальному головастику, и это говорило о том, что из двух половинок раннего эмбриона земноводного могут появиться два совершенно одинаковых животных.
Но когда Шпеман разделил эмбрион по-другому, перевязав его перпендикулярно области контакта между двумя первыми клетками, эффект был совершенно иным. С одной стороны образовывался нормальный головастик, а с другой стороны разрасталась неорганизованная масса ткани. В конечном итоге это помогло установить, что участок эмбриона, называемый дорсальной губой бластопора, играет ключевую роль в развитии эмбриона. Если этот участок удалить, эмбрион превращается в сгусток ткани, в котором отсутствуют структуры, в норме формирующие спинную (дорсальную) сторону туловища. Кроме того, если дорсальную губу бластопора пересадить на будущую брюшную (вентральную) сторону другого эмбриона, происходит образование второй эмбриональной оси и развиваются два сросшихся эмбриона (рис. 2.2). Шпеман назвал этот участок "организатором", поскольку установил, что он организует образование нервных тканей в дорсальной части эмбриона и может инициировать развитие дополнительной оси тела при пересадке[4].
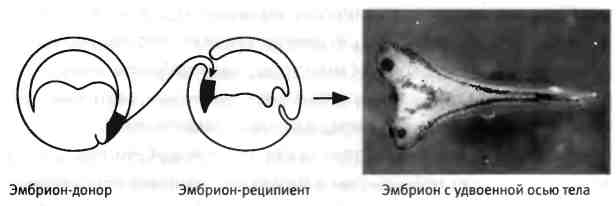
Рис. 2.2. Индукция дополнительной (вторичной) оси тела. Пересадка ткани "организатора" в другой участок эмбриона вызывает образование еще одного эмбриона, "сиамского близнеца", соединенного с первым. Фотографию предоставили Хироки Курода и Эдди де Робертис, Калифорнийский университет в Лос-Анджелесе.
Впечатляющий эксперимент Шпемана показал, что упорядоченное развитие эмбриона обеспечивается взаимодействием между его частями. Были обнаружены и другие организаторы с такими же поразительными свойствами, показавшие, как этот принцип работает на самых разных уровнях: на уровне целого эмбриона, отдельных его частей и вплоть до мельчайших структур. Давайте познакомимся с двумя другими организаторами, не менее важными, чем дорсальная губа бластопора.
Образование конечностей всегда вызывало восторг у эмбриологов. Начиная с маленького бугорка на боку у зародыша, конечность проходит в своем развитии множество стадий, пока не примет окончательную форму. У трехдневного эмбриона цыпленка этот бугорок составляет в длину и в ширину не более 1 мм, но к моменту вылупления из яйца конечность увеличивается в размере примерно в тысячу раз. В ходе развития маленькая лапка растет и удлиняется, приобретает кости, хрящи, мускулы, сухожилия, пальцы и перья, прекрасно демонстрируя скоординированность процессов развития. Возможно, самым удивительным является строго упорядоченный процесс образования хрящевой ткани (которая впоследствии заменяется костной тканью). Хрящ образуется вокруг плотного скопления клеток и закладывается по направлению от плеча к кисти и далее к пальцам. За ходом процесса можно проследить с помощью специальных красителей (рис. 2.3). Строгий порядок событий при развитии конечностей и наличие у конечности полярности, которую мы наблюдаем в виде четкой последовательности пальцев, означают, что зачаток конечности, как и эмбрион в целом, может каким-то способом проинструктировать клетки, чем они должны в конце концов стать.

Рис. 2.3. Формирование конечностей у цыпленка. Всего за несколько дней в ходе эмбрионального развития ноги и крылья цыпленка очень сильно увеличиваются. За образованием хрящевой ткани, предшествующим формированию костей, можно проследить с помощью специальных красителей Этот процесс начинается с проксимальных частей конечностей и заканчивается формированием пальцев. Обратите внимание, что детали анатомического строения крыльев и ног различаются. Фотографии предоставлены Джозефом Лэнкманом и Джоном Фоллоном, факультет анатомии Университета Висконсина.
Несколько десятилетий назад другой пионер эмбриологии, Джон Сондерс, открыл в почке крыла куриного эмбриона организатор полярности конечности. В норме куриное крыло имеет три пальца, которые по размеру и форме можно идентифицировать как пальцы 2, 3 и 4 (в направлении от передней к задней части крыла; пальцы 1 и 5 в крыле не формируются). Когда Сондерс пересадил кусочек ткани из задней части зачатка крыла (оттуда, где должен был появиться 4-й палец) в переднюю часть крыла (где в норме появляется 2-й палец), сформировалось крыло, имеющее дополнительные пальцы. Эти дополнительные пальцы представляли собой зеркальное отображение нормальных пальцев: вместо последовательности пальцев 2, 3, 4 возникла последовательность 4, 3, 2, 3, 4 (рис. 2.4). Такая зеркальная полярность говорит о том, что клетки в задней зоне почки крыла организуют полярность последовательности пальцев (4, 3, 2), и если эти клетки пересадить в другое место, точно такая же последовательность возникает и там.
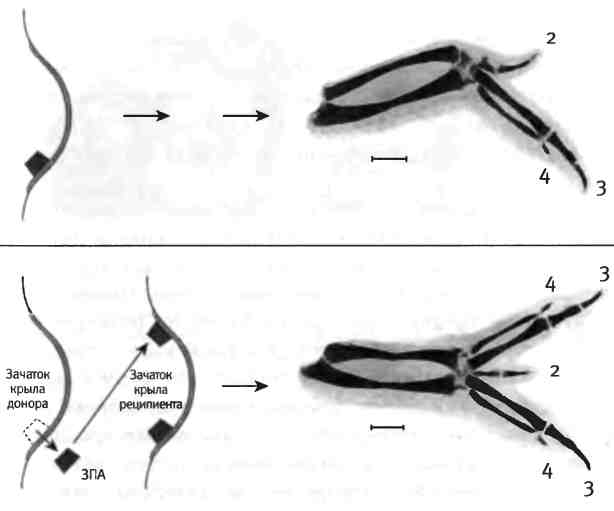
Рис. 2.4. Индукция полидактилии у курицы. Трансплантация зоны поляризующей активности (ЗПА) из заднего участка зачатка развивающегося крыла в новое место в передней зоне приводит к появлению дополнительных пальцев, последовательность которых имеет полярность, противоположную нормальной. Фотографии предоставлены Джозефом Лэнкманом и Джоном Фоллоном, факультет анатомии Университета Висконсина.
Сферы влияния организатора Шпемана и зоны поляризующей активности (ЗПА) в конечности цыпленка достаточно широки. Эти организаторы воздействуют на развитие всего эмбриона или значительных участков его тела. Однако были обнаружены организаторы, действующие на развивающиеся структуры гораздо меньшего размера. В 1980 г. Фредерик Нийхут из Университета Дьюка показал, что формирование пятен-глазков на крыльях бабочек также индуцируется организаторами. Когда Нийхут убивал микроскопическую группу клеток, которая должна была формировать центр глазка, глазок не образовывался. Более того, он обнаружил, что если эту группу клеток удалить из крыла бабочки в первые сутки развития на стадии куколки и пересадить в любой другой участок крыла, пятно-глазок появляется на новом месте (рис. 2.5). Причем такой способностью обладают только клетки из центральной части пятна. Нийхут назвал организатор пятна-глазка "фокусом".

Рис. 2.5. Индукция развития пятна-глазка на крыле бабочки. Пересадка клеток из центральной части глазка в развивающемся крыле в другое место приводит к появлению глазка на этом участке. Фотография Фредерика Нийхута из его книги "Развитие и эволюция рисунка на крыльях бабочек" (The Development and Evolution of Butterfly Wing Patterns).
Все организаторы обладают способностью влиять на морфогенез (процесс, приводящий к созданию формы) тканей или клеток. Такую их специфическую активность объясняют в первую очередь тем, что клетки организаторов производят вещества, влияющие на развитие других клеток. Такие вещества называются морфогенами. Влияние организаторов зависит от их расстояния до клеток-мишеней: клетки, расположенные поблизости от организатора, испытывают наиболее сильное воздействие, а те, что расположены дальше в эмбрионе тритона, зачатке конечности или крыле бабочки, подвергаются меньшему воздействию. Уже давно появилось предположение о том, что морфогены, образующиеся в клетках определенного участка, диффундируют оттуда, создавая градиент концентрации. То есть идея такова, что клетки, окружающие источник морфогена, реагируют на него в соответствии с полученным количеством вещества. Например, в почке крыла курицы клетки, расположенные вблизи ЗПА, формируют задний палец (палец 4), а те, что расположены дальше, формируют следующие пальцы (соответственно, 3 и 2). В пятнах-глазках на крыльях бабочек концентрические круги окрашенных в разные цвета чешуек, по-видимому, являются результатом ответа на уровень морфогена, синтезируемого клетками "фокуса", разный в зависимости от расстояния от источника.
Морфогены, ответственные за активность организаторов, можно назвать Чашей Грааля в эмбриологии. Основной трудностью в этой области исследований было то, что активность организаторов является результатом действия целой группы клеток. Любая клетка производит тысячи веществ, и нельзя было исключать, что за активность организаторов отвечают несколько субстанций. Пересадка клеток — это мощный инструмент исследования, но эмбриологам предстояло отыскать способ находить морфогены в густом биохимическом бульоне. Ждать пришлось несколько десятилетий.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК