Сборка позвоночных: больше Hox-генов и много сдвигов
Эволюцию нашей семейной линии тоже можно проследить до самого кембрия. Мы относимся к позвоночным животным, которые, в свою очередь, принадлежат к хордовым — животным, обладающим хордой. Кроме позвоночных, к хордовым также относятся оболочники (такие как асцидии) и головохордовые (такие как ланцетники). Хордовые — вторичноротые животные (рис. 6.8). Долгое время наиболее изученным древним хордовым животным оставалась Pikaia из отложений в Берджесских сланцах, однако недавние находки в Чэнцзяне показали, что позвоночные существовали еще раньше, примерно 520 млн лет назад, причем их анатомия была на удивление сложной.
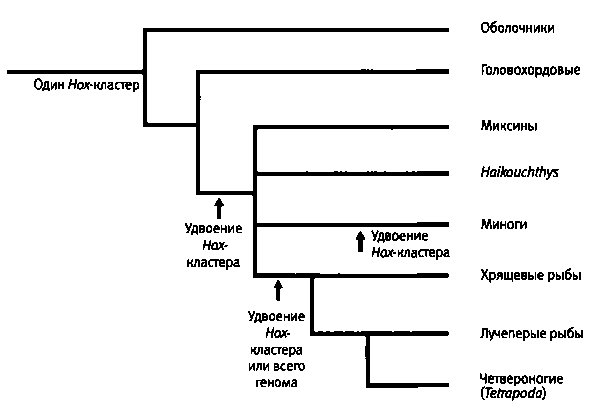
Рис. 6.8. Эволюционное древо хордовых животных и увеличение числа кластеров Hox-генов в ходе эволюции позвоночных. Общий предшественник всех хордовых имел один кластер генов, как и ныне живущие оболочники и головохордовые. Удвоение кластера генов в ходе эволюции происходило несколько раз — в линии бесчелюстных рыб, в линии хрящевых рыб (акул), а также у миног. Поскольку эволюционное происхождение кембрийского позвоночного Haikouichthys точно неизвестно, его ветвь принято изображать отходящей от древа одновременно с ветвями миксиновых рыб, миног и хрящевых рыб. Рисунок Джоша Клейса.
Образцы ископаемой бесчелюстной рыбы Haikouichthys ercaicunensis демонстрируют наличие головной лопасти с глазами, возможно, носовые отверстия, десять или более отдельных позвонков, жабры, а также спинной и брюшной плавник. Такое строение намного сложнее, чем у более позднего вида Pikaia, что говорит о том, что в начале кембрийского периода эволюция позвоночных животных продвинулась уже достаточно далеко. Эти недавние открытия подчеркивают чрезвычайную важность поиска ископаемых животных в уже известных и в еще не исследованных отложениях. Датировка первого появления той или иной группы животных или тех или иных признаков всегда очень примерная, поскольку последующие находки могут отодвинуть ее назад. В данном случае новые находки позволили отодвинуть происхождение группы на 15 млн лет назад. Кроме того, хотя в начале и середине кембрийского периода позвоночные не были самой многочисленной группой животных, их обнаружение в Чэнцзяне позволяет вписать хищника Haikouichthys в кембрийскую экосистему.
Появление позвоночных сопровождалось изобретением и модификацией многих структур и систем тела, включая более сложный мозг, органы чувств, хрящи, скелет и череп. Множество последующих эволюционных новшеств привело к формированию пресмыкающихся, земноводных, птиц и млекопитающих, которых мы видим сегодня. Как и в случае членистоногих и лопастеногих, нам хотелось бы знать, определялась ли ранняя эволюция позвоночных в кембрийском периоде тем же набором генов развития, которым обладали и другие группы животных, или же происхождение предковых позвоночных связано с какими-то изменениями в наборе генов развития.
Мы не можем изучить гены Haikouichthys, однако мы можем проанализировать гены некоторых современных животных, занимающих ключевые позиции на филогенетическом древе хордовых и вторичноротых, что позволит оценить генетическую сложность древних позвоночных. Одна из ключевых групп — головохордовые. У этих животных нет таких признаков позвоночных как череп или скелет, но они являются сестринской группой по отношению к позвоночным, точно так же как современные онихофоры являются сестринской группой по отношению к членистоногим. Набор генов Hox-кластера у головохордовых должен соответствовать набору генов Hox-кластера последнего общего предшественника головохордовых и позвоночных.
Единственным ныне живущим представителем головохордовых является ланцетник. Это животное длиной 5-8 см можно встретить в заливе Тампа во Флориде и в некоторых других местах. Когда Джорди Гарсия-Фернандес и Питер Холланд впервые занялись изучением его Hox-генов, они обнаружили всего один Hox-кластер. Вспомните, что современные позвоночные, такие как мышь и человек, имеют по четыре кластера, в сумме содержащих 39 генов. Изучение генов ланцетника позволяет сделать вывод о том, что увеличение числа кластеров Hox-генов произошло уже после разделения линий позвоночных и головохордовых (в кембрийском периоде или несколько раньше). Мы также знаем, что другие вторичноротые, такие как оболочники и иглокожие, обладают единственным кластером Hox-reнов. Значит, в то время как все без исключения оболочники и иглокожие в кембрийском периоде и позднее эволюционировали при наличии единственного кластера, содержащего десять Hox-генов, позвоночные увеличили число Hox-генов.
Когда в ходе эволюции позвоночных увеличилось число кластеров Hox-генов? Стало ли это событие толчком к эволюции позвоночных? Чтобы ответить на эти вопросы, пришлось изучить множество современных видов, относящихся к разным группам на разных ветвях филогенетического древа позвоночных. Все млекопитающие, птицы и некоторые рыбы, включая древнего глубоководного целаканта, имеют по четыре кластера Hox-генов. У нас есть все основания заключить, что у общего предка этих челюстных позвоночных также было четыре кластера Hox-генов.
Однако более примитивные современные позвоночные, такие как минога, имеют меньшее число Hox-кластеров. Детальное изучение Hox-кластеров миноги и их сравнение с Hox-кластерами костистых рыб и млекопитающих показало, что наши четыре кластера — результат двух раундов дупликации на ранних этапах эволюции позвоночных. Первое удвоение случилось после расхождения головохордовых и предков миног, а второе — за некоторое время до появления костистых рыб. Посмотрим еще раз на наше семейное древо (рис. 6.8). Поскольку образцы из Чэнцзяня являются бесчелюстными рыбами, мы можем предположить, что они имели всего один или два кластера Hox-генов.
Выясняется, что различие в числе кластеров Hox-генов у позвоночных отражает различие в общем количестве генов развития у этих животных. В ходе эволюции позвоночных произошло удвоение не только Hox-кластера, но и многих других генов развития. Это могло произойти, например, при дупликации всего генома или его крупного фрагмента. Увеличение числа генов развития у высших позвоночных говорит о том, что на ранних этапах эволюции позвоночных появление новых генов сыграло значительную роль в эволюции плана строения. Одним из показателей эволюции анатомии хордовых является количество клеток разных типов. Так вот, у человека и других высших позвоночных гораздо больше типов клеток, чем у головохордовых: у этих последних нет клеток, из которых у нас состоят хрящи, кости, голова и некоторые органы чувств. Это означает, что большее количество генов развития коррелирует с большим количеством клеточных типов и более сложной организацией тканей, поскольку большее количество генов позволяет генерировать большее количество инструкций в ходе эмбрионального развития.
Однако главной движущей силой последующей эволюции высших позвоночных было не увеличение количества генов. Важно помнить, что эволюция земноводных, пресмыкающихся, птиц и млекопитающих происходила при наличии все тех же четырех кластеров Hox-генов. Лягушки и змеи, динозавры и страусы, жирафы и киты эволюционировали с одним и тем же набором Hox-генов. Таким образом, общее число Hox-генов ничего не говорит нам об эволюции форм животных. Разнообразное строение этих видов, как и в случае членистоногих, стало результатом изменения географии эмбрионов за счет сдвига зон экспрессии Hox-генов, а не результатом увеличения числа этих генов.
Например, у позвоночных переход от одного типа позвонков к другому — шейные/грудные, грудные/поясничные, поясничные/крестцовые, крестцовые/хвостовые — соответствует границам между зонами экспрессии определенных Hox-генов. Передняя граница экспрессии одного из генов, Hoxc6, совпадает с границей между шейным и грудным отделами у мыши, курицы и гуся, хотя у всех этих животных разное количество шейных позвонков. Таким образом, у этих животных произошло смещение зоны экспрессии гена Hoxc6 по отношению к номеру позвонка (рис. 6.9). У змей это смещение выражено еще сильнее. У них нет четкой границы между шейным и грудным отделами позвоночника, и экспрессия Hoxc6 продолжается до самой головы. Ко всем этим позвонкам присоединены ребра, а это означает, что все эти позвонки грудного типа, однако при этом у них есть еще и признаки шейных позвонков. Отсюда следует, что удлинение тела змеи произошло за счет потери шеи и удлинения грудного отдела в результате сдвига зон экспрессии Hox-генов.
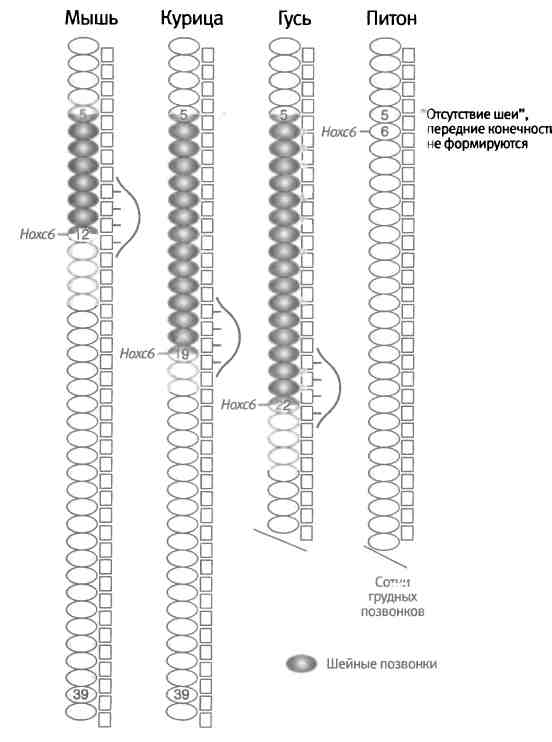
Рис. 6.9. Сдвиг зон экспрессии Hox-генов также способствует эволюции позвоночных. Разные группы позвоночных животных отличаются друг от друга количеством шейных позвонков: у мыши шея короткая, у гуся длинная, а у питонов шеи практически нет, а есть просто длинное туловище. Граница между шейными и грудными позвонками у всех позвоночных совпадает с границей экспрессии гена Hoxc9, однако у разных животных это положение относительно всего тела различается. На этом уровне у всех четвероногих позвоночных возникают передние конечности, а у змей эта граница сдвинута вперед к основанию черепа и конечности не развиваются. Рисунок Лианн Олдс.
Одновременно удивительно и логично, что эволюция формы тела представителей двух наиболее успешных и разнообразных групп животных — членистоногих и позвоночных — происходила по одному и тому же сценарию, связанному со сдвигом зон экспрессии генов Hox вдоль основной оси эмбриона. Самое главное здесь — что мы ухватили суть крупномасштабных изменений в строении животных. Теперь мы можем рассматривать отдельные группы животных — насекомых, пауков и многоножек или птиц, млекопитающих и пресмыкающихся, а также их давно вымерших предков — не как нечто уникальное, а как вариации на общую тему. Лучше всего эту мысль сумел выразить блестящий писатель и философ второй половины восемнадцатого века Дени Дидро (см. цитату в начале главы), и ее же почти сто лет назад сформулировал в своем законе Сэмюель Уиллистон. Теперь мы знаем общий механизм и, следовательно, очень точное объяснение одной из важнейших тенденций в эволюции животных.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК