Летаем и ползаем: эволюция конечностей для нового образа жизни
За последующие 350 млн лет строение и функции конечностей четвероногих животных менялись множество раз во многих направлениях — от удивительного превращения лап в крылья у летающих животных до различной степени редукции конечностей у обитателей воды и суши. Все эти модификации происходили за счет эволюционных изменений развития конечностей, и в некоторых случаях ученым, работающим в области эво-дево, удавалось зафиксировать некоторые важные сдвиги в географии развивающихся конечностей.
Три раза в ходе эволюции животных — у птерозавров, птиц и летучих мышей — передние конечности тетрапод превращались в крылья. Чтобы животное могло летать, крыло должно двигаться вверх и вниз, вперед и назад, а также складываться вдоль тела, когда животное находится на земле. Интересно, что дизайн крыльев каждой из трех групп летающих животных отличается важными деталями. Пэт Шипман в книге "Расправляя крылья" (Taking Wing) назвала крылья птерозавров "крыльями пальцев", крылья птиц "крыльями предплечий", а крылья летучих мышей "крыльями кистей" (рис. 7.10). Давайте рассмотрим эти три изобретения в порядке их появления, начиная с птерозавров.
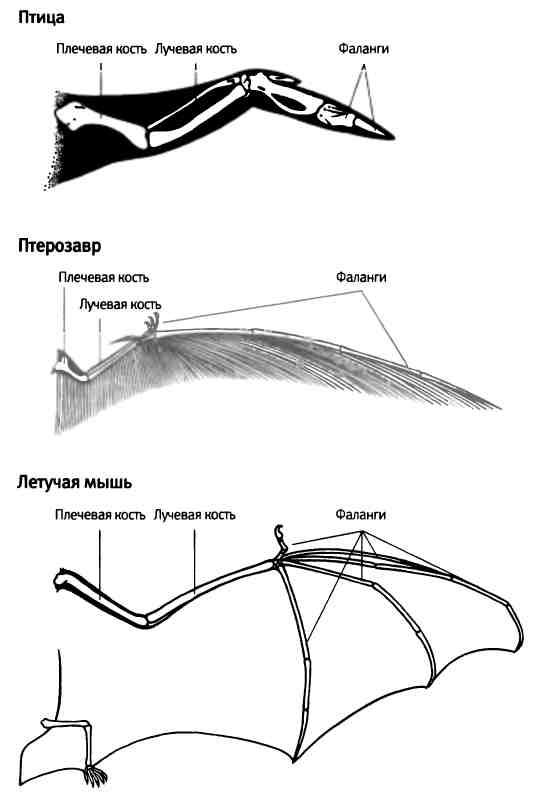
Рис. 7.10. Эволюция крыльев позвоночных животных. Крылья птиц — это "крылья предплечий", на которых перья растут вдоль всей длины конечности. Крылья птерозавров — "крылья пальцев": большая часть перепонки крыла связана с длинным пальцем. Крыло летучей мыши — "крыло кисти", поскольку поверхность крыла соединена со многими пальцами передней конечности и простирается до задней конечности. Рисунок Лианн Олдс.
Птерозавры поднялись в воздух примерно 227 млн лет назад — за 70 млн лет до того, как появились птицы (птицы берут свое начало от оперенных динозавров, а не от птерозавров). Самой яркой отличительной чертой крыльев птерозавров был очень длинный четвертый палец, составлявший основу внешней части крыла. Эта конечность имела все основные элементы структуры, включая три первых пальца, а кости кисти были слиты воедино. Три первых пальца не были связаны с перепонкой крыла. Перепонка распространялась на всю длину конечности, но основной размах крыла создавался за счет вытянутого четвертого пальца.
У птиц крыло не имеет перепонки, а состоит из перьев, которые представляют собой выросты кожи по всей длине конечности. Основная часть поверхности крыла приходится на предплечье, тогда как вклад плечевой кости и кисти с пальцами значительно меньше. На самом деле все четыре пальца птиц очень короткие.
Крыло летучей мыши представляет собой кожистую мембрану, опирающуюся на предплечье и сильно вытянутые пальцы, со 2-го по 5-й, так что такое крыло можно назвать "крылом кисти". Задний край крыла достает до задней конечности и прикрепляется на уровне пятки; это придает животному дополнительную устойчивость при полете.
Различия в архитектуре крыльев отражают модификации развития на основе базовой структуры передней конечности тетрапод. Нам многое известно о сходстве в формировании передних конечностей птиц и млекопитающих, однако некоторые детали, определяющие различия крыльев птиц и летучих мышей, пока неясны и активно изучаются биологами-эволюционистами.
О некоторых сдвигах мы знаем больше из модификации развития конечностей у змей и некоторых видов рыб. У змей произошло удлинение всего тела, а развитие конечностей оказалось полностью подавленным. У питонов и удавов по-прежнему образуются рудименты задних конечностей, но передние отсутствуют полностью. В принципе, подавление развития конечностей может происходить на нескольких этапах — от стадий, предшествующих образованию почки конечности, до более поздних стадий ее формирования.
Изучение развития конечностей у эмбриона питона позволило обнаружить эволюционные изменения ранних этапов формирования почки конечности, ответственные за безногость. В частности, распространение зон экспрессии некоторых Hox-генов на всю длину туловища животного до самой головы привело к исчезновению участков инициации формирования почек передних конечностей. Зачатки задних конечностей по-прежнему образуются, но их рост прерывается. Прекращение развития задних конечностей коррелирует с отсутствием экспрессии ключевых сигнальных белков, включая белок Sonic hedgehog, в регионе-организаторе почки задней конечности. Около анального отверстия на туловище питонов и удавов действительно образуются рудименты задних конечностей (анальные шпоры), но у представителей более молодых семейств змей нет даже этих рудиментов. По-видимому, полное отсутствие конечностей у этих змей является результатом нарушения развития конечностей на еще более раннем этапе.
Эволюция конечностей ни в коем случае не сводится к этим давнишним событиям в истории основных групп позвоночных животных. Адаптация и эволюция конечностей продолжаются до сих пор, и у некоторых молодых видов (в частности, у саламандр и ящериц) такие параметры, как число пальцев, варьируют в достаточно широких пределах. Эволюция плавников рыб также является очень динамичным процессом. Недавняя модификация скелета трехиглой колюшки способствовала тому, что этот вид рыб стал одной из излюбленных моделей для изучения эволюции скелета позвоночных животных. Во многих озерах северной части Северной Америки обитают две формы колюшки, относительно недавно произошедшие от общего морского предка. Когда в конце последнего ледникового периода, примерно 15 000 лет назад, ледники начали отступать, популяции колюшек остались жить в ледяных озерах. Затем на протяжении короткого (в геологическом плане) промежутка времени эти популяции дали начало двум формам: первая обитает в придонных слоях шельфовой зоны и имеет короткие шипы, а вторая живет на больших глубинах и отличается значительно большей длиной шипов на спине и на животе (рис. 7.11).
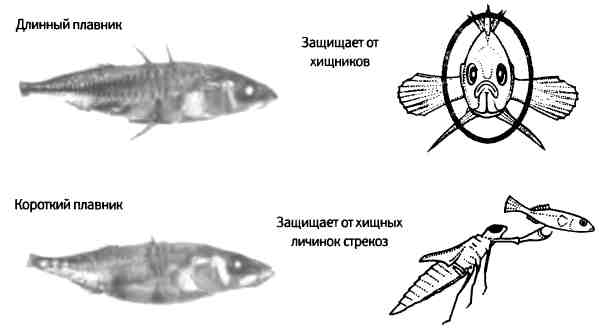
7.11. Эволюция скелета трехиглой колюшки. Длинные шипы защищают колюшку, обитающую на большой глубине, от нападения хищников, поскольку увеличивают общий диаметр туловища. Однако для колюшки, обитающей на мелководье, эти шипы становятся недостатком, так как за них хватаются хищные личинки стрекоз, поэтому сокращение длины этого плавника позволяет ускользнуть от хищников. Рисунок предоставлен Дэвидом Кингсли, Медицинский институт Говарда Хьюза и Стэнфордский университет.
Эти две формы в особенности различаются средствами защиты, состоящими из жестких костных пластинок по бокам и шипов, торчащих из верхней и нижней части тела. Количество и длина этих шипов коррелируют с активностью хищников в водоеме. В открытых водоемах более длинные шипы защищают колюшку от пасти хищника. Однако на дне длинный брюшной плавник становится недостатком. Как это ни парадоксально, серьезная угроза для колюшки — личинка стрекозы, которая хватает молоденьких рыб за плавники. В условиях такого сильного давления хищников в естественных популяциях несколько раз быстро эволюционировала форма, лишенная шипов.
Брюшной плавник является задней конечностью рыбы, так что его укорочение происходит за счет изменений в процессе развития задних конечностей. Специалистам в области биологии развития удалось многое узнать о генах, вовлеченных в развитие и дифференцировку передних и задних конечностей. Один из этих генов, Pitxi, участвует в формировании задних конечностей у тетрапод и брюшного плавника у рыб. Анализ экспрессии Pitxi у не имеющей шипов колюшки из озер Британской Колумбии показал, что экспрессия этого гена в зачатках брюшных плавников специфическим образом отсутствует. Это эволюционное изменение регуляции гена Pitx1, как вы уже могли догадаться, связано с изменением переключателя этого гена, предотвращающим его экспрессию в задних конечностях. Эволюционные изменения переключателя позволили изменить функцию гена Pitxi в брюшном плавнике, но не повлияли на функции этого гена в других областях эмбриона.
Кроме того, анализ генов колюшки с редуцированными шипами, на этот раз из Исландии, говорит о том, что у этих рыб независимым образом произошло такое же изменение экспрессии гена Pitxi. Уменьшение размера шипов также хорошо документировано у ископаемых видов, причем отнюдь не близкородственных. Это говорит о том, что сокращение размера брюшного шипа — достаточно распространенное явление, связанное с многократными эволюционными изменениями переключателей гена Pitxi. Эти наблюдения, а также приведенный в предыдущей главе пример эволюции ногочелюстей показывают, что некоторые эволюционные изменения не являются редкими единичными событиями, но происходят в различных популяциях и у различных видов при одинаковом давлении отбора. В этом смысле можно сказать, что эволюция "воспроизводима".
Действительно, редукция плавника или конечности — совсем не редкое явление. У двух далеких групп млекопитающих — китообразных (киты и дельфины) и ламантинов — произошло значительное сокращение размеров задних конечностей в ходе их независимой эволюции от сухопутных предков к полностью водному образу жизни. Эволюция безногих ящериц также происходила многократно. Таким образом, колюшка — не какое-то странное исключение, но хорошая модель для изучения достаточно распространенного и важного эволюционного процесса. Кроме того, эти рыбы прекрасно сохранились в виде окаменелостей, возраст которых превышает много тысячелетий. Анализ этих окаменелостей показывает, что сокращение размеров плавников может происходить менее чем за 10 000 поколений, или меньше чем за 10 000 лет. Конечно, это не одно мгновение, но на геологической шкале это очень короткий интервал. Подробная палеонтологическая летопись и хорошо изученная генетика современных популяций колюшки, а также повторяющиеся независимые примеры одинаковых эволюционных изменений делают колюшку одним из наиболее выразительных примеров эволюции.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК