Отбор, гены и приспособленность: каким должно быть преимущество?
В двух последних главах я привел аргументы и свидетельства в пользу того, что пятна-глазки на крыльях бабочек, а также окраска кошек-меланистов, темных и светлых мешотчатых прыгунов и полосатых зебр эволюционировали под действием естественного отбора, а пятна на крыльях дрозофил сформировались в ходе полового отбора. В некоторых случаях очевидно, что те или иные признаки дают своим носителям определенные преимущества. Однако насколько велико должно быть преимущество, чтобы отбор благоприятствовал носителю того или иного признака? В 1910 году Тедди Рузвельт не увидел, чтобы пятна леопарда или полоски зебры давали своим хозяевам какие-либо преимущества. Я подозреваю, что Рузвельт, как и многие другие люди, считал, что преимущество должно быть очевидным или измеряемым и что лишь тогда оно приводит к отбору соответствующего признака. Итак, вопрос звучит следующим образом: насколько велико должно быть различие, чтобы оно имело значение?
Здесь мы внедряемся в сферу популяционной генетики — раздела генетики, который исследует изменчивость особей в популяции, генетические основы такой изменчивости и изменения частоты, с какой те или иные фенотипические формы и гены встречаются в ходе эволюции. Коротко ответить на вопрос, насколько велико должно быть различие, можно так: для того, чтобы заработал естественный отбор, достаточно на удивление малого различия в успешности двух форм. Это различие часто невозможно увидеть или измерить в полевых условиях, но его достаточно, чтобы в ходе эволюции одна из форм получила преимущество над другой.
Популяционные генетики вывели несколько формул, которые выявляют основные связи между преимуществами или недостатками определенной мутации и судьбой этой мутации внутри популяции или вида. С помощью этих формул можно рассчитать, насколько одна фенотипическая форма должна быть "лучше" другой, чтобы распространиться в популяции. И — как много времени это займет.
Тут нужно иметь в виду несколько факторов и понятий. Когда мы говорим "лучше", что мы на самом деле имеем в виду? Существует понятие "приспособленность": для животных приспособленность складывается из выживаемости (продолжительности жизни отдельной особи) и плодовитости (количества потомков). Чтобы естественный отбор сработал и новая мутация закрепилась в популяции, она должна давать относительное преимущество в приспособленности. Допустим, что особи с новой мутацией (например, меланизм у мотыльков или мешотчатых прыгунов) оставляют в среднем по 101 потомку, тогда как особи без этой мутации оставляют лишь по 100 потомков. В этом случае различие в относительной приспособленности составляет 1%. Для использования в нашей формуле мы преобразуем это значение в коэффициент отбора (s), который в данном случае составляет 0,01.
Имеет ли значение столь слабое различие? Если преимущество в 1% сохранится, то да. Частота встречаемости данной мутации в популяции будет возрастать со скоростью, которая зависит от размера популяции и значения коэффициента отбора. Для расчета времени распространения мутации в популяции, измеряемого в числе поколений, пользуются следующей формулой:
Время = 2 / s x ln(2N)
Здесь N — число особей в популяции, a In — натуральный логарифм.
В нашем примере, если N = 10 000, что является достаточно большим и вполне реалистичным числом, a s = 0,01, для распространения мутации в популяции потребуется (2/0,01) In (2 х 10 000) = 1980 поколений. Для мышей или мотыльков это соответствует примерно 2000 годам или менее. Если s = 0,001, т. е. преимущество составляет лишь 0,1%, мутация все-таки закрепится в популяции, но только через 20 000 поколений. Эти расчеты показывают, что даже очень небольшое преимущество позволяет мутации распространиться в популяции за достаточно короткий срок (по геологической шкале). Однако коэффициент отбора вовсе не обязательно низкий. Быстрое повышение частоты встречаемости мотыльков-меланистов в промышленных районах Англии или насекомых, устойчивых к инсектицидам, происходило за считаные годы, а не за тысячелетия. В этих случаях коэффициенты отбора находятся в интервале от 0,2 до 0,5, то есть являются довольно высокими и отражают огромные селективные преимущества.
Обсуждая возможности естественного отбора, нельзя не учитывать и обратный процесс — удаление из популяции неблагоприятных мутаций. Я не буду углубляться в арифметику, лишь замечу, что мутации, которые вызывают даже очень незначительный недостаток, почти не имеют шанса распространиться в большой популяции. Обсуждая преимущества и недостатки темного и светлого меха у мешотчатых прыгунов, стоит задуматься о том, что каких-то вариантов рисунка мы в природе не встречаем — например, не существует пятнистых мешотчатых прыгунов. Может быть, такая мутация в принципе невозможна? Я думаю иначе. Если такие животные появляются, они вынуждены жить либо на темной, либо на светлой почве, и в любом случае их окраска является настолько невыгодной, что в дикой природе они не могут достичь значительной численности.
В конце этой главы л предлагаю вернуться к спору о зебре, используя ту же логику, с помощью которой я только что объяснил отсутствие пятнистых мышей. Подумайте, ведь все зебры на свете полосатые. Если бы полосы не давали преимущества, разве мы не встречали бы множество зебр без полосок? Конечно, встречали бы. Мутации окраски у млекопитающих происходят достаточно часто, так что в дикой природе с очень небольшой частотой встречаются и белые тигры, и пятнистые зебры. Заводчики, занимающиеся разведением домашних животных, на протяжении многих веков отбирали редкие спонтанно возникавшие варианты, например, разнообразные варианты окраски лошадей — родственников зебр. Я думаю, что лаборатория африканской саванны доказывает нам, что полоски действительно имеют значение.
Мы просто не знаем, для какой цели они нужны. Вспомните теорию: минимальное преимущество перед теми, у кого полос нет, — это все, что нужно, чтобы полосы сохранялись. Базовый принцип действия естественного отбора (включая половой отбор) по созданию или сохранению какого-либо признака распространяется на эволюцию всех видов, включая нас с вами. То же самое относится к основным закономерностям эво-дево, касающимся модульного строения, генетических переключателей и эволюции формы. В следующей главе мы, наконец, поговорим о том, как появился Homo sapiens и его специфические признаки.
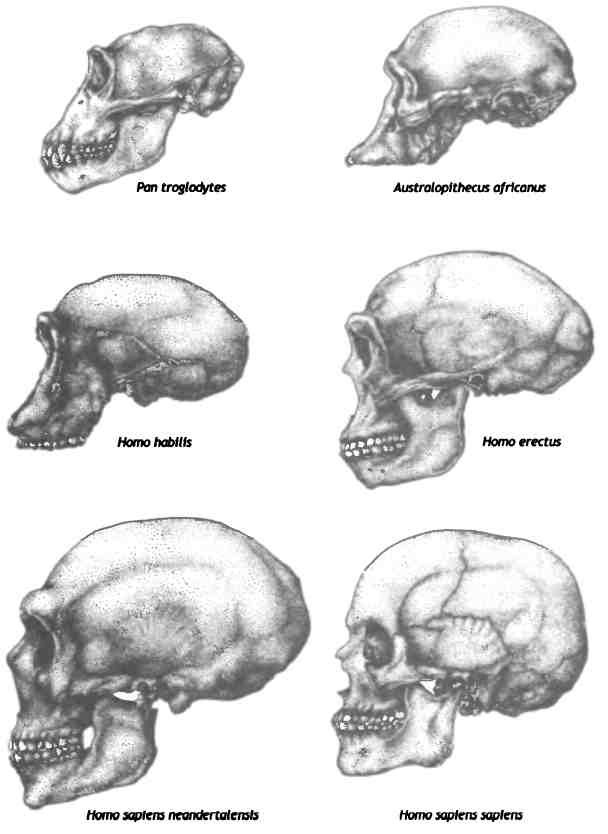
Эволюция формы и размера черепа гоминид. Рисунок Деборы Майзелс, Zoobotanica.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК