Сдвиг зон экспрессии Hох-генов и закон Уиллистона
Эволюция членистоногих в кембрийском периоде главным образом состояла в увеличении числа и разнообразия сегментов и конечностей. Тело трилобита состоит из трех основных отделов — головы, туловища и пигидия (хвостового отдела), причем сегменты каждого из этих отделов и отходящие от них конечности очень похожи друг на друга и различаются только по размеру. У ныне существующих групп членистоногих, представители которых появились не позднее чем через 150 млн лет после окончания кембрийского периода, конечности гораздо разнообразнее — их насчитывается более десяти видов. Отростки на голове, туловище и хвосте являются специализированными структурами, которые служат для питания, восприятия сигналов, передвижения, дыхания, рытья нор, копуляции, заботы о потомстве и защиты. Успешное развитие членистоногих, безусловно, связано с их способностью к адаптации, которая, в свою очередь, объясняется возрастающей специализацией конечностей.
Но как возникли разные типы конечностей? По всей видимости, этот процесс должен был сопровождаться серьезными изменениями географии эмбрионов членистоногих. Чтобы понять, что конкретно произошло в ходе эволюции членистоногих, мы вновь обратимся к анализу современных животных. Лучше всего генетический контроль дифференцировки конечностей изучен у дрозофил. Нам известно, что образование каждого типа конечностей (разнообразных отростков на голове, каждой из трех пар ног, обычно отсутствующих конечностей на брюшке, а также гениталий, которые также представляют собой модифицированные конечности) контролируется Hox-белками. Разнообразие формы и функций конечностей достигается путем экспрессии разных Hox-генов в разных участках тела вдоль главных осей эмбриона. При формировании эмбриона образуется множество зон экспрессии Hox-генов по отдельности и в разных сочетаниях (на рис. 6.7 эти зоны обозначены цифрами от 1 до 10).
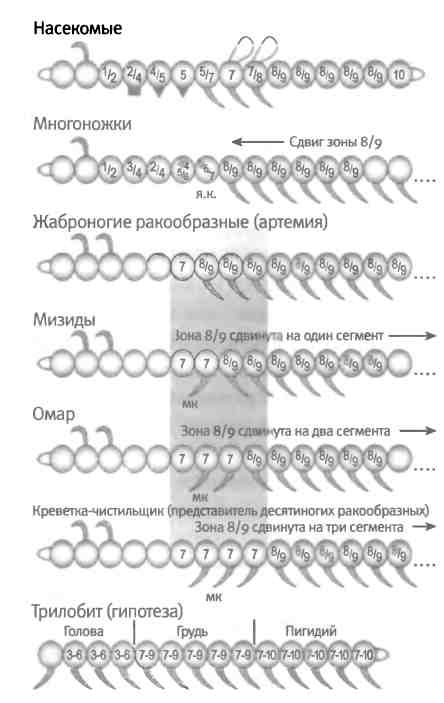
Рис. 6.7. Сдвиг зон экспрессии Hox-генов определяет основные различия в форме тела членистоногих. Hox-гены обозначены номерами. Обратите внимание на сдвиг границы зоны экспрессии генов 7, 8 и 9 у насекомых, многоножек и четырех ракообразных (артемий, мизид, омаров и креветок-чистильщиков) (затемненная область). Количество ногочелюстей, или максилл (мк), находится в обратной зависимости от количества сегментов, в которых экспрессируются гены 8/9, и сдвигается в сторону увеличения от варианта артемий, у которой нет ногочелюстей. У многоножки непосредственно перед ногами располагается ядовитый коготь (я. к). У трилобитов, по-видимому, было всего три отдела тела, различающихся тремя разными комбинациями зон экспрессии Hox-генов. Рисунок Лианн Олдс.
Каким было строение эмбрионов кембрийских животных? Как экспрессировались их Hox-гены 500 млн лет назад? Увидеть этого мы не можем, но можем сделать некоторые выводы, сравнивая географию эмбрионов и картину экспрессии Hox-генов у разных ныне живущих членистоногих. Например, некоторые членистоногие, такие как жаброногие рачки, имеют очень простое строение грудного отдела, все сегменты и конечности которого очень похожи. По-видимому, таким же строением обладал их примитивный предок. На рис. 6.7 показано, что у эмбрионов жаброногих ракообразных зоны экспрессии двух Hox-белков (номера 8 и 9) идентичны, тогда как у насекомых зоны их экспрессии различаются. У многоножек, еще одной представительной группы членистоногих, рисунок экспрессии Hox-генов в эмбрионе напоминает таковой у жаброногих. Длинное туловище состоит из идентичных сегментов, несущих одинаковые конечности. У эмбриона многоножки те же два Hox-белка (номера 8 и 9) экспрессируются в каждом из сегментов и в отходящих от них конечностях. У этих двух членистоногих зоны идентичных сегментов соответствуют зонам экспрессии одного и того же (или одних и тех же) Hox-белков. Таким образом, можно сделать вывод, что у членистоногих кембрийского периода, таких как трилобиты, блоки похожих сегментов и отростков соответствовали зонам экспрессии одних и тех же Hox-белков.
Нам также известно, что границы между Hox-зонами у членистоногих обычно соответствуют смене типов сегментов и конечностей. У жаброногих и многоножек в сегменте, расположенном непосредственно перед грудным отделом, экспрессируется уже другой Hox-белок или комбинация Hox-белков (номера 7 и 5/6/7 соответственно) и образуются конечности другого типа. У жаброногих это конечности ротового аппарата, а у многоножки эта конечность стала ядовитым когтем, необходимым для иммобилизации жертвы и для защиты. Такая связь между различными типами конечностей и различными Hox-зонами, расположенными вдоль оси эмбриона, распространена достаточно широко.
Известно, что у членистоногих смещение зон экспрессии Hox-белков тесно коррелирует с эволюционными различиями в количестве и форме конечностей, развивающихся на соответствующих сегментах. Причем подобные различия наблюдаются не только между представителями основных классов членистоногих, но и внутри классов. Роль сдвига зон экспрессии генов Hox в эволюции прекрасно продемонстрировали Михалис Авероф и Нипам Пател, которые собрали и изучили эмбрионы широкой выборки ракообразных (к этой группе членистоногих относятся креветки, усоногие раки, крабы и омары). Одним из заметных различий между группами было разное количество ногочелюстей (максилл), представляющих собой модифицированные конечности и расположенных в передней части грудного отдела. У жаброногих ракообразных ногочелюстей нет, нет их и у примитивных ракообразных. Однако более сложные ракообразные имеют одну, две или даже три (как омар) пары максилл. За эти важные различия отвечают небольшие изменения в географии эмбрионов. Авероф и Пател обнаружили, что у обладателей ногочелюстей зоны экспрессии двух Hox-белков (номера 8 и 9) сдвинуты назад соответственно на один, два и три сегмента по сравнению с ракообразными, у которых ногочелюсти отсутствуют (рис. 6.7). Величина сдвига точно коррелирует с количеством ногочелюстей. Более того, по-видимому, этот сдвиг и образование ногочелюстей в ходе эволюции происходили у ракообразных несколько раз независимым образом, что говорит о том, что сходные функциональные адаптации у разных животных достигаются за счет использования одного и того же механизма. Подробнее о значении повторяющихся изменений я расскажу в следующей главе.
Сдвиг зон экспрессии Hox-генов стал причиной выраженных различий тела по главной оси таких групп современных членистоногих, как пауки, ракообразные, многоножки и насекомые. С большой долей вероятности можно предположить, что то же самое происходило и в кембрийском периоде, поскольку регионализация тела и специализация конечностей наблюдаются у всех ископаемых членистоногих того времени. Блоки сходных сегментов у разных видов ископаемых членистоногих определенно соответствовали зонам экспрессии конкретных Hox-генов (рис. 6.7). Возникновение новых типов конечностей и сегментов в эволюции членистоногих связано с появлением у их эмбрионов большего числа уникальных зон экспрессии отдельных Hox-генов и их комбинаций. Таким образом, относительный сдвиг зон экспрессии Hox-генов является одним из механизмов, лежащих в основе закона Уиллистона: для специализации повторяющихся частей тела требуется, чтобы они находились в зонах экспрессии разных Hox-генов.
Сдвиг зон экспрессии Hox-генов происходил не только у членистоногих. Тот же механизм лежит в основе формирования анатомического разнообразия и нашей с вами ветви — позвоночных.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК