Глава 14 Аналитическая и экспериментальная эмбриология
Эмбриология последней четверти XIX в. характеризуется появлением новых проблем. Описательная эмбриология предшествующего периода была в состоянии ответить на вопрос «как» — проследить, иногда вдаваясь в очень тонкие детали, последовательность изменений отдельных частей и органов развивающегося зародыша. Сравнительно-описательный метод использовался, кроме того, для выяснения филогенетических отношений. Однако такой путь исследования не давал возможности ответить на вопрос «почему», т. е. выяснить условия и вскрыть причины отдельных процессов, из которых складывается сложная картина эмбрионального развития. Здесь нужны были другие методы — описательно-аналитический и, особенно, экспериментальный.
Путь анализа и эксперимента, приводящий к расчленению сложных явлений на составляющие их частные процессы, давно уже завоевал права гражданства в науках о неорганической природе — физике и химии, а среди биологических наук — в физиологии. В науке о наследственности — генетике — этот путь был намечен в исследованиях Г. Менделя. Для изучения индивидуального развития, в котором особенно отчетливо обнаруживается гармоничная целостность организма, аналитический и экспериментальный методы первоначально казались неприменимыми. Считалось, что расчленение морфологического целого на составляющие его компоненты должно привести к исчезновению самого существенного, чем характеризуются живые организмы в отличие от тел неживой природы.
Эти скептические и совершенно лишенные основания соображения высказывались до недавнего времени даже тогда, когда на путях аналитического и экспериментального изучения эмбрионального и постэмбрионального развития были достигнуты существенные успехи, когда этими методами удалось выяснить новые, ранее неизвестные стороны формообразовательных процессов в онтогенезе. Попытки теоретического обобщения полученных таким образом результатов действительно натолкнулись на трудности, которые не удавалось преодолеть на том методологическом уровне науки об индивидуальном развитии, на котором исследователям представлялся выбор лишь между преформистским механистическим и эпигенетическим виталистическим взглядами на живую природу.
«Органообразующие участки зародыша».
В 1874 г. вышла небольшая книга немецкого анатома и эмбриолога В. Гиса «Форма нашего тела и физиологическая проблема ее возникновения». Появлением этой книги датируется начало аналитического направления в эмбриологии. Гис утверждал, что топография органов позднего зародыша и развившегося из него сформированного организма является следствием закономерного расположения зачатков этих органов на ранних эмбриональных стадиях.

Вильгельм Гис. 1831–1904.
Имея в виду зародышевый диск куриного зародыша, Гис писал: «С одной стороны, каждая тонка в эмбриональной области зародышевого диска должна соответствовать будущему органу или части органа, и, с другой стороны, каждый орган, возникающий из зародышевого диска, имеет предобразованную закладку в каком-либо пространственно определимом участке плоского диска… Материал для закладки существует уже в плоском зародышевом диске, но морфологически он не отчленен и потому как таковой непосредственно не обнаруживается. На пути ретроспективного прослеживания мы должны прийти к тому, чтобы и в периоде, когда морфологическое расчленение неполно или когда его нет совсем пространственно определить место каждой закладки; желая быть последовательными, мы должны распространить это определение и на только что оплодотворенное яйцо. Принцип, согласно которому зародышевый диск содержит предобразованные в распластанном виде закладки органов… я называю принципом органообразующих участков зародыша… Все закладки, имеющиеся в зародышевом диске, растут, но их рост происходит без соответствия с первоначальным отношением размеров; одни растут быстрее, другие медленнее, одни перестают расти раньше, другие позже, и в результате того, что каждая из них растет в меру присущего ей особого закона, будущие органы отличаются от первичных закладок не только по взаимному расположению, но и по относительной массе. Этот важный принцип мы называем принципом неравномерного роста»[104].
Сложные формообразовательные процессы Гис стремился объяснить неравномерным ростом, изгибаниями клеточных слоев зародыша, а в конечном счете сводил все эти изменения к игре механических сил. В сентябре 1867 г. А.О. Ковалевский в письме к И.И. Мечникову сообщал, что проездом из Парижа в Триест он был в Базеле у Гиса и видел его препараты. К теоретическим выводам Гиса Ковалевский отнесся с осторожностью: «Он (Гис) старается все процессы объяснить механически, и много весьма удачных выводов, но и увлечения также немало; у него введены даже формулы, сделаны модели из воску, и разные приспособления он объясняет при посредстве гуттаперчевых трубок и пластинок»[105].
Основное положение Гиса, основанное на аналитическом описании раннего эмбрионального развития, как следует из приведенной выше выдержки из его книги, сводится к представлению о преформированных в бластодерме органообразовательных участках. К этому же положению, хотя и не в столь категорической форме, пришли исследователи, использовавшие в эмбриологии принципиально новый метод — экспериментальный.
Каузально-аналитический метод.
Возникновение этого нового направления в эмбриологии связано с исследованиями и теоретическими взглядами немецкого анатома и эмбриолога В. Ру. Ему принадлежит первая удачная попытка активного воздействия на ход эмбрионального развития, имевшая целью причинный анализ процессов формирования зародыша.

Вильгельм Ру. 1850–1924.
По мнению Ру, причины, управляющие индивидуальным развитием, не могут быть выяснены простым описанием; здесь необходимо «каузальное, следовательно, аналитическое мышление», которое, таким образом, объединяет декларированное Ру экспериментальное направление в учении о развитии индивидуума с аналитическими исследованиями, основывающимися на описательном методе, в частности, с исследованиями Гиса.
Применение экспериментального метода к изучению процессов индивидуального развития заключается в том, что сложная организованная система развивающегося организма и условия среды, необходимые для осуществления развития, расчленяются на отдельные компоненты; Ру назвал эти компоненты факторами, или причинами процесса развития. Такое расчленение позволяет, по мнению Ру, выяснить характер участия каждого из этих компонентов в развитии и в реализации определенного результата этого процесса. Выключение того или иного фактора из развивающейся системы посредством оперативного удаления какой-либо ее части или искусственное устранение действия какого-либо фактора окружающей среды позволяет выяснить зависимость направления развития от данного фактора. Обратный опыт — включение в систему ранее элиминированного внешнего или внутреннего фактора — дает возможность проверить результаты первого опыта. Ру считал, что при помощи подобных аналитических экспериментов можно установить различие между участвующими в развитии факторами и разделить их на две основные группы — определяющих, или детерминирующих, с одной, стороны, и реализующих, или индифферентных, с другой.
«Детерминирующие» и «индифферентные» факторы развития.
Согласно представлениям Ру, детерминирующими факторами являются такие компоненты организма или окружающей его среды, от которых зависит направление развития, качественно-специфические особенности его результата. В отличие от них, реализующие факторы необходимы лишь для обеспечения самого хода процессов развития, направление и исход которых от них не зависят.
В соответствии с этим, одна из важных задач науки, которую Ру назвал каузальной морфологией, или механикой развития, состоит, по его мысли, в выяснении локализации определяющего фактора. Если каузально-аналитический эксперимент показывает, что фактор, от которого зависит направление развития всего зародыша или его части, локализован в самой живой системе (в развивающемся зародыше или определенной его части), следует говорить об автономном дифференцировании, или самодифференцировании. Если же, наоборот, обнаруживается, что детерминирующий фактор находится вне системы, т. е. во внешней по отношению ко всему зародышу среде (а по отношению к его части — или вне организма, или внутри него, но в другой его части), то такой случай следует называть зависимым дифференцированием.
Ру считал, что наряду с выяснением локализации определяющих факторов (внутри развивающейся системы или вне ее) в задачи механики развития входит установление времени начала действия фактора, длительности и интенсивности его действия, а также способов действия определяющих факторов на детерминируемый ими процесс развития. Ру видел задачу механики развития в обнаружении комплексных факторов индивидуального развития. Разложение их на простые факторы, идентичные тем, которые участвуют в явлениях неорганического мира, он считал преждевременным. По мнению Ру, сведения биологических закономерностей к законам физики и химии может рассматриваться как реальная, но отдаленная перспектива дальнейшей работы.
Материализм Ру имеет, таким образом, все признаки непоследовательного, механистического материализма. Логическим следствием такого хода мысли часто оказывались идеалистические представления о нематериальном целедеятельном принципе, который принимался за источник целостности сложного организма. Сам Ру против этой формы идеализма в биологии, получившей название неовитализма, решительно возражал. Он полемизировал с теоретиком неовитализма Г. Дришем и считал совершенно недопустимым введение в науку таких понятий, как энтелехия[106].
Основные положения механики развития, которые декларировал, применял в собственной научной работе и неустанно пропагандировал Ру в течение почти полустолетия, легли в основу длинного ряда экспериментальных исследований его современников.
Мозаичная теория развития.
Исследования, легшие в основу этой теории, открываются классическим опытом Ру на яйце лягушки. Желая выяснить вопрос, предопределено ли развитие частей зародыша в только что начавшем дробиться яйце, Ру разрушал нагретой иглой один из первых двух бластомеров. При этом он обнаружил, что бластомер, оставшийся неповрежденным, дает начало половинному зародышу с одним нервным валиком, зачатком глаза и слуховым пузырьком. Отсюда Ру сделал вывод, что в первых двух бластомерах преформированы правая и левая половины будущего зародыша.
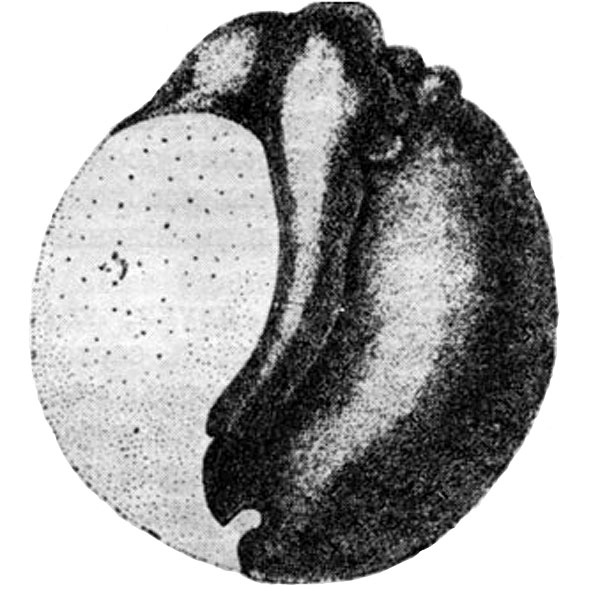
Половинный зародыш, полученный разрушением одного из двух первых бластомеров лягушки (по В. Ру, 1881).
Основываясь на описанных опытах, Ру сформулировал так называемую «мозаичную теорию» развития. Он писал, что развивающаяся гаструла лягушки и формирующийся из нее зародыш, начиная со второго дробления — мозаичное образование, состоящее по крайней мере из четырех вертикальных, независимо развивающихся частей.
Сходные результаты одновременно и независимо от Ру получил бельгийский зоолог Л. Шабри на начальных стадиях дробления асцидии. Шабри пришел к заключению, что каждый бластомер содержит потенции определенных частей и что его разрушение оказывается невосполнимой утратой. Следовательно, различные части тела животного преформированы в различных участках яйца. Иными словами, данные Ру и Шабри служили как бы экспериментальным подтверждением положения Гиса об «органообразовательных участках», явившегося предшественником мозаичной теории эмбрионального развития.
Указанная теория созвучна представлениям А. Вейсмана о дифференцировании зародыша в результате неравнонаследственного деления ядра. Развивая свою умозрительную теорию наследственности, Вейсман предполагал, что гипотетические носители наследственных свойств (детерминанты) яйцевого ядра распределяются при каждом дроблении неравномерно, что и лежит в основе процессов дифференцирования зародыша.
Явление мозаичности в развитии было обнаружено различными авторами также в опытах с разделением первых бластомеров у развивающихся гребневиков (Г. Дриш и Т. Морган, Г. Циглер, Н. Ятсу) и моллюсков (Г. Кремптон и Э. Вильсон, Э. Конклин). Впрочем, довольно скоро выяснилось, что этот вывод во многих отношениях неверен. Прежде всего сам Ру убедился, что в ряде случаев при разрушении одного бластомера образуется не только хорошо сформированная половина зародыша, но и в различной степени другая его половина. Это явление Ру назвал «постгенерацией». Оно противоречило представлению, согласно которому в правом бластомере заложены материалы только для правой, а в левом — только для левой половины тела.
«Мозаичные» и «регуляционные» яйца.
Еще более противоречили этому представлению результаты опытов с отделением друг от друга первых бластомеров у кишечнополостных, немертин, ланцетника и амфибий. Бластомеры яйца морского ежа, помещенного в морскую воду, лишенную солей кальция, при легком встряхивании разделяются и сохраняют способность развиваться дальше. В опытах такого рода Г. Дриш (1891) показал, что при делении яиц на стадии двух или четырех бластомеров из каждого бластомера образуется целый зародыш, размеры которого соответственно в два или в четыре раза меньше, чем у нормального. Вильсон получил тот же результат на яйцах ланцетника, а Р. Зойа (1895) в опытах на яйцах медуз установил, что целые зародыши получаются при изоляции бластомеров даже на 16-клеточной стадии. А. Герличка (1896) и Г. Шпеман (1901–1903) разделяли два первых бластомера у тритона волосяной петлей и получали в ряде случаев из каждого бластомера целый жизнеспособный зародыш. Если перетяжка не полностью отделяла бластомеры друг от друга, развивались сращенные и в различной степени обособленные двойниковые уроды.
Одним из важных следствий опытов Шпемана было опровержение гипотезы Вейсмана о неравнонаследственном делении при дроблении яйца как источнике дифференцирования. Шпеман накладывал волосяную петлю, не затягивая ее, на оплодотворенное, но еще не начавшее дробиться яйцо тритона так, чтобы яйцевое ядро оказывалось в одной половине получившейся гантелеобразной фигуры. В результате дробилась только та часть яйца, в которой находилось ядро. После нескольких дроблений лигатура расслаблялась, и это давало возможность одному из ядер проскользнуть в недробившуюся половину яйца. Лигатура затягивалась снова, и каждая половина яйца продолжала развиваться самостоятельно. Несмотря на то, что в половине яйца, начавшей дробиться раньше, оказывались, например, 7/8 материала первоначального яйцевого ядра, а в другой половине — только 1/8, из обеих половин яйца развивались полноценные зародыши.
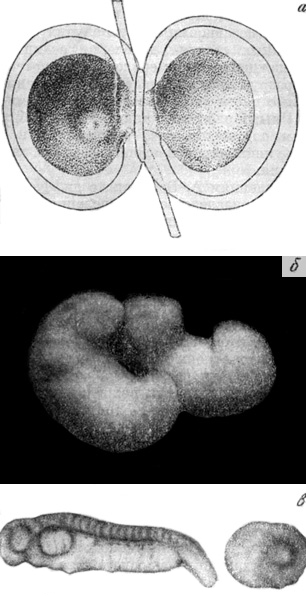
Опыт Шпемана.
а — наложение волосяной петли на оплодотворенное яйцо тритона; б — результат разделения яйца в медианной плоскости — двойниковые зародыши; в — результат разделения яйца во фронтальной плоскости: зародыш и недифференцированный «брюшной кусок».
Из сопоставления мозаичного раннего развития аспидий и моллюсков с регуляционным развитием медуз, ланцетника и амфибий первоначально был сделан вывод, что одни животные характеризуются мозаичными, а другие — регуляционными яйцами. Однако на многих объектах, прежде всего на яйцах морского ежа (Т. Бовери, 1901), удалось показать, что развитие первоначально имеет «регуляционный» характер, а затем делается «мозаичным». В течение первых двух дроблений составные части протоплазмы яйца, именно слои, пересекаемые под прямым углом вертикальной осью яйца, распределяются между четырьмя бластомерами поровну, при изоляции этих бластомеров каждый из них, естественно, может дать начало целому зародышу. В результате третьего дробления из образовавшихся восьми бластомеров верхняя четверка получает одни протоплазменные материалы, а нижняя — другие; поэтому при изоляции бластомеров на 8-клеточной стадии каждый из них уже не в состоянии образовать целый, обладающий всеми частями организм.
Проблемы формообразования в индивидуальном развитии сделались ареной столкновения механистического и виталистического взгляда на живую природу.
Механистическая и виталистическая концепции эмбрионального развития.
Мозаичной теории развития, имевшей отчетливо выраженный механистический характер, Дриш противопоставил виталистические взгляды, к которым он пришел, доведя до логического конца механистическое представление о развивающемся организме. Аналитическое расчленение целого эмбриологами-экспериментаторами привело Дриша (1908) к мысли о существовании иррационального остатка анализа — непознаваемого, внепространственного целедеятельного фактора (энтелехии), которому он приписывал свойство определять целостность и целесообразные свойства развивающегося зародыша. Эта разновидность витализма (энтелехиальный витализм) явилась следствием банкротства механистического понимания жизненных явлений.
Эволюция воззрений Дриша, одного из виднейших представителей неовитализма конца XIX и начала XX в., чрезвычайно поучительна. Дриш начал свою научную деятельность как исследователь, давший блестящие образцы применения экспериментального метода к изучению явлений эмбриональных и постэмбриональных регуляций. Помимо получения интересных фактических данных, Дриш пришел в своих работах к важным теоретическим обобщениям. Так, он сформулировал положение, что в развивающемся организме проспективная потенция его частей шире, чем их проспективное значение. Это означает, что отдельная часть организма (а равно и организм в целом) может реализовать не только ту возможность (проспективное значение), которая осуществилась при данных условиях, но и другие возможности (в сумме названные проспективной потенцией), для осуществления которых должны, конечно, быть созданы соответствующие условия. Приведенное обобщение Дриша послужило теоретической основой направленного воздействия на развивающийся организм, т. е. основой как в экспериментальном методе при лабораторном изучении формообразования, так и для изменения направления развития в практических целях. Дриш, в сущности, подвел итог исследованиям многих ботаников и зоологов, доказывавших пластичность форм и возможность их экспериментального изменения. Новую область биологии, занимающуюся экспериментальным изучением развития организма, Ру, как это было отмечено выше, называл механикой развития, Дриш — физиологией развития, К.А. Тимирязев — экспериментальной морфологией, а М.М. Завадовский — динамикой развития.

Ганс Дриш. 1867–1941.
В отличие от механистов, допускавших строгую детерминацию частей развивающегося организма и предопределение их будущей судьбы в развитии, виталисты, в частности Дриш, считали способный к регуляционным изменениям организм «гармонично-эквипотенциальной системой», так как, по их мнению, части этой системы первоначально обладают равными возможностями (потенциями), а итогом развития оказывается гармонично построенное целое. Проспективное значение части развивающегося целого (В), по Дришу, может быть выражено формулой B=f(S,l,E), которая выражает функциональную зависимость проспективного значения части от ее величины B, положения в целом l и присущей целому организму потенции вызывать формирование данной части в том или ином направлении; это свойство целого — энтелехия Е. Ей, как считал Дриш, может быть дано только отрицательное определение: энтелехия не энергия, не сила, не константа, не субстанция; она не связана с чем бы то ни было материальным; она внепространственна и вообще может мыслиться только как понятие. Определение энтелехии, данное Дришем в книге «Философия органического» (1908), достаточно ясно показывает, что это ненаучное понятие. Вместе с тем конкретные исследования Ру, Дришем и другими эмбриологами-экспериментаторами явлений формообразования наметили тот путь, следуя которому в первой половине XX в. эмбриология смогла обогатиться большим количеством фундаментальных открытий.
Маркировка раннего зародыша.
Аналитическое и экспериментальное изучение индивидуального развития, успешно начатое в 80-х годах XIX в., широко развернулось за порогом XX столетия.
Выяснение закономерностей превращения частей оплодотворенного яйца или зародыша на стадии бластулы в органы и системы сформированного организма осуществлялось разными методами. Один из них, связанный с именем немецкого эмбриолога В. Фогта, заключался в нанесении красочных меток на отдельные участки поверхности бластулы амфибий и в наблюдении за перемещением этих меток. Таким образом удалось установить, что одни метки на последующих стадиях, прежде всего во время гаструляции, оставались на поверхности, входя в состав покровов и закладки нервной системы, другие погружались в глубь зародыша, образуя закладки хорды и осевой мезодермы.

Вальтер Фогт. 1888–1941.
Перемещения клеток и клеточных комплексов, изменяющие конфигурацию частей зародыша, все более приближающуюся к окончательной, а также качественные изменения отдельных клеток и их групп — дифференцирование, изучались как аналитическим, так и экспериментальным путем.
Теория поля.
Советский биолог А.Г. Гурвич сделал смелую попытку подойти к анализу формообразовательных перемещений отдельных клеток, клеточных комплексов или слоев, опираясь на теорию поля. Наряду с исследованием морфогенеза у грибов и цветковых растений Гурвич изучал в свете этой теории также эмбриональный морфогенез центральной нервной системы и закладку скелета конечностей позвоночных. Согласно его первоначальной гипотезе, абстрагированной от гистологического строения какой-либо развивающейся части организма, источником направленного роста закладки является «поле», действие которого ограничивает формативные движения ее элементов, подобно тому, как магнитное поле ориентирует железные опилки. В последующих работах Гурвича, объединенных в 1944 г. в книге «Теория биологического поля», эта концепция вылилась в представление об элементарных анизотропных клеточных полях, синтезирующихся в поле целого (эмбриональной закладки, органа или организма) по правилам сложения векторов.
Понятие «поля» в сочетании с понятием «градиент» широко использовалось в специальных работах и сводках, посвященных общим проблемам формообразования в индивидуальном развитии (Г. Шпеман, 1936; Дж. Хаксли и Г. де Вер, 1932; К.X. Уоддингтон, 1957; и др.). Теория поля, в понимании Гурвича, продолжает разрабатываться Л.В. Белоусовым, которому принадлежит, в частности, исторический очерк развития этой теории (1963).
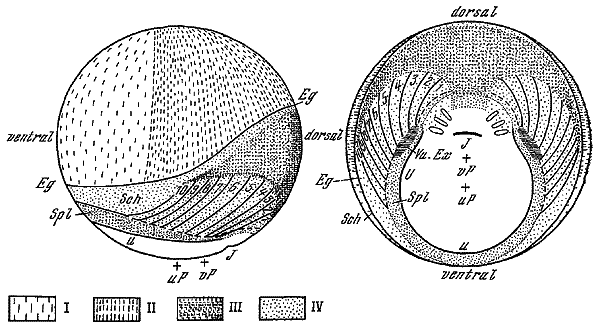
Карта распределения презумптивных зачатков в бластуле тритона, полученная с помощью витальных меток (по В. Фогту, 1925).
I — эпидермис; II— нейтральная пластинка; III — хорда; IV — мезодерма; 1-10 — будущие мезодермальные сегменты.
Элементарные модусы формообразования. Дифференцирующие деления.
К области аналитического изучения онтогенеза относятся также исследования, посвященные закономерностям клеточного и тканевого дифференцирования и роста. Изменение формы зародыша в целом и его частей не может быть сведено к одному элементарному процессу, в частности, к изменению взаимного положения или формы клеток. Например, при инвагинационной гаструляции происходит не просто впячивание одной стенки шарообразной бластулы, но и характерные внутриклеточные структурные и функциональные изменения.
Среди элементарных способов, или модусов формообразования наиболее сложным для изучения является именно дифференцирование, источником которого являются как влияние на клетки извне — воздействие соседних клеток и окружающей среды, так и внутренние процессы в самих клетках, выражающиеся в появлении в результате деления материнской клетки двух неодинаковых дочерних клеток. Следовательно, в первом случае речь идет о дифференцирующем взаимодействии, изучавшемся преимущественно экспериментальным путем, а во втором — о дифференцирующих делениях.
Попытку объяснить дифференцирование в онтогенезе неравномерным распределением составных частей клеток при их делении впервые сделал в конце XIX в. А. Вейсман. Его идея о дифференциальном, неравнонаследственном делении клеточного ядра, как уже было сказано, не подтвердилась, однако представление о существовании дифференцирующих делений возродилось впоследствии, когда было с несомненностью установлено, что различия в судьбе каждой из двух дочерних клеток могут возникать в результате самого деления произведшей их материнской клетки. К числу явлений этого рода относится прежде всего неравномерное распределение материалов протоплазмы дробящегося яйца между отдельными бластомерами.
Дифференцирующие деления на более поздних стадиях развития особенно подробно описывались у насекомых. Обзор явлений этого рода приведен в книге А. Кюна «Лекции по физиологии развития» (1955). Дифференцирующие деления описаны, в частности, при развитии фасетчатых глаз у муравьев и при образовании крыловых чешуек у бабочек.
Примеров, в которых дифференцирующий характер клеточных делений был бы точно доказан цитологическими описаниями, немного. Существует, однако, огромное количество явлений дифференцирования и на преэмбриональных этапах (образование гамет), и на всех стадиях эмбрионального и постэмбрионального развития, которые с очень большой долей вероятности могут быть объяснены наличием делений; в результате этих делений дочерние клетки оказываются с самого момента их возникновения не тождественными друг другу.
Анализ явлений роста.
Среди явлений индивидуального развития аналитическими методами наиболее успешно исследовались процессы роста. При аналитическом изучении роста увеличение размеров целого организма или отдельных его частей рассматривалось прежде всего с чисто количественной стороны, что выражалось в попытке найти математическое выражение скорости этого процесса. Предлагались эмпирические кривые и соответствующие им уравнения, подобранные по принципу наибольшего совпадения со сглаженной кривой, получаемой непосредственным измерением линейных, объемных или весовых данных, относящихся к растущим организмам или их частям.
Другой способ получения уравнений роста основывался на определенных теоретических предпосылках, лежащих в основе понимания сущности явлений роста. Т.Б. Робертсон (1923) исходил из представления о росте как процессе, совершающемся неограниченно долгое время всей массой организма или органа, и выражал его уравнением, идентичным уравнению скорости мономолекулярной аутокаталитической реакции. Эта формула роста подверглась справедливой критике, поскольку давно уже было известно, что относительная скорость роста с возрастом обычно убывает. Ч. Майнот (1913) высказал предположение, что это замедление зависит от прогрессирующего дифференцирования клеток, постепенно утрачивающих вследствие этого способность к размножению. Из этого же предположения исходили авторы теорий роста, независимо друг от друга разрабатывавшие эту проблему — С. Броди в США, Дж. Хаксли в Англии и И.И. Шмальгаузен в СССР — и пришедшие к выводу, что зависимость между размерами зачатка, органа или целого организма и временем может быть выражена уравнением параболы. Изучение изменений скорости роста органов при развитии куриного зародыша в сопоставлении с литературными данными, относящимися к росту рыб и человека, позволили Шмальгаузену сделать обобщение, что такой параболический рост типичен для всех случаев, когда рост сопровождается прогрессивным дифференцированием. Шмальгаузен (1935) показал, что процесс роста может быть охарактеризован константой k, являющейся произведением удельной скорости роста C? и возраста t:
С? ? t = k.
Удельная скорость роста представляет бесконечно малое приращение за бесконечно малый отрезок времени, отнесенное к единице размера, т. е.
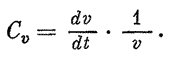
Величина k остается постоянной в течение каждого обособленного периода жизни. При переходе от одного периода жизни к следующему величина константы роста обычно уменьшается, что является выражением обратной зависимости между ростом и дифференцированием.
Выражение процессов роста уравнением параболы дает возможность математического описания отношений роста между отдельными частями тела или между частью и целым организмом, т. е. так называемого относительного роста.
Методы экспериментальной эмбриологии.
В XX в., особенно начиная с 20-х годов, широко развернулись экспериментальные исследования эмбрионального развития. Стремление преодолеть виталистическое истолкование эпигенеза, связанное, главным образом, с воззрением Дриша, вылилось в попытки представить формообразовательные процессы в онтогенезе как результат взаимного влияния частей развивающегося зародыша. Основателем этого направления в эмбриологии по справедливости считается немецкий эмбриолог Г. Шпеман, разработавший тонкие методы хирургического вмешательства в процессы эмбрионального развития.
Объектом исследования Шпемана, его учеников и многочисленных последователей были преимущественно зародыши амфибий на ранних стадиях развития. Используя простые инструменты — тонко отточенные скальпели, микропипетки, волосяные петли и стеклянные иглы, — эмбриологи шпемановской школы успешно осуществляли удаление определенных, точно локализованных участков зародыша и пересадки таких участков с одного места на другое того же или другого зародыша.
Если на стадии бластулы или ранней гаструлы пересадить участок будущей спинной эктодермы одного зародыша на брюшную или боковую сторону другого, то пересаженный участок развивается не так, как он развивался бы, будучи оставлен на месте. Этот участок дает начало не той или иной области головного или спинного мозга, а, ассимилируясь окружающей покровной эктодермой, принимает участие в образовании кожи. Другими словами, пересаженный участок эктодермы развивается не в соответствии с его происхождением, а в соответствии с местом, где он оказывается после операции. Равным образом, участок будущей покровной эктодермы, взятый с брюшной или боковой области бластулы или ранней гаструлы и пересаженный на спинную сторону, превращается не в кожу, которая должна была бы развиться на месте, откуда этот участок был изъят, а в часть нервной системы хозяина. Достоверность этих результатов обеспечивалась, в частности, применением гетеро- или ксенотрансплантации, т. е. пересадкой упомянутых участков от одного вида хвостатых амфибий другому, например, от зародыша гребенчатого тритона зародышу альпийского, или даже пересадкой в пределах разных отрядов амфибий, например, от зародыша бесхвостой амфибии — жерлянки зародышу хвостатой амфибии — тритону. Различия клеток трансплантата и клеток хозяина позволяли с уверенностью судить, какие части химерного зародыша развились из пересаженного материала и какие — из тканей хозяина под влиянием трансплантата.
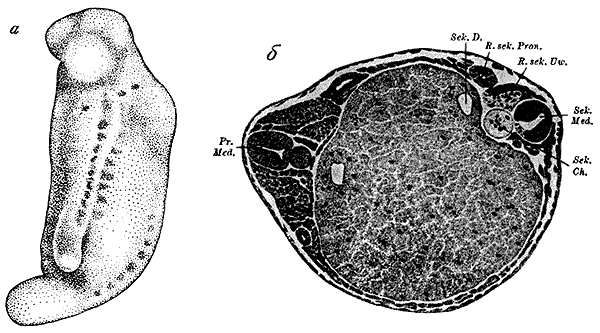
Индукция вторичного зародыша тритона (по Г. Шпеману и Г. Мангольд, 1924).
а — зародыш, на боку которого развился индуцированный зародыш со слуховыми пузырьками, медуллярной трубкой и двумя рядами сомитов; б — поперечный разрез через зародыш с первичными (слева) и вторичными (справа) индуцированными осевыми органами.
Результат получается иной, если описанные операции провести на стадии поздней гаструлы. В этом случае участок спинной эктодермы образует на брюшной или боковой стороне включенную в кожу часть центральной нервной системы, а участок брюшной или боковой эктодермы, пересаженной на спинную сторону, дает в центральной нервной системе начало тканям кожи. Иначе говоря, а этой стадии пересаженные на другое место участки зародыша развиваются уже не в соответствии с новым местоположением, а в соответствии с их происхождением. Из этих опытов был сделан вывод, что при переходе к стадии поздней гаструлы у зародыша утрачивается способность изменять направление развития под влиянием нового окружения и, наоборот, закрепляется способность развиваться в одном определенном направлении.
Детерминация. Открытие первичного организатора.
Это закрепление формативных свойств эмбрионального материала получило название детерминации. Источником детерминации частей развивающегося организма Шпеман считал влияние одной части зародыша на другую.
Зависимость развития одной части зародыша от другой Шпеман открыл задолго до того, как было обнаружено упомянутое явление детерминации. В 1901 г. он сообщил об опытах удаления глазной чаши у зародышей травяной лягушки на стадии, когда закладки хрусталика, развивающегося из покрывающей глазную чашу эктодермы, еще нет; в отсутствие глазной чаши хрусталик, а равно и роговица, не развиваются. Наоборот, если пересадить зачаток глаза под эктодерму любой части тела, или, если участок туловищной эктодермы пересадить поверх глазного зачатка, то из эктодермы, в норме не образующей хрусталика и роговицы, при контакте с глазной чашей развиваются хрусталик и прозрачная роговица.
Применительно к развитию осевых органов (осевой скелет, мускулатура, центральная нервная система) детерминационное взаимодействие частей доказывалось следующим образом. Во время гаструляции происходит впячивание эмбрионального материала, лежащего в определенной области поздней бластулы; этот материал, как показывают опыты прижизненной красочной маркировки соответствующих участков зародыша, подстилает изнутри спинную эктодерму, дающую начало нервной системе, и образует так называемую крышу первичной кишки, средняя часть которой является будущей хордой: ее боковые мезодермальные участки развиваются в дальнейшем в туловищные мышцы, осевой скелет и выделительные органы. В процессе гаструляции этот погружающийся в глубь зародыша материал постепенно проходит через пограничный участок между анимальной и вегетативной половинами зародыша, ограничивающий сверху отверстие бластопора, т. е. через верхнюю, или спинную, губу бластопора. Если вырезать на стадии поздней гаструлы кусочек верхней губы бластопора и пересадить его другому, несколько более молодому зародышу в бластоцель (полость бластулы или ранней гаструлы), то из трансплантата на новом месте разовьются хорда и органы, в норме образующиеся из осевой мезодермы (мышцы, скелет и каналы предпочки). Под влиянием этого трансплантата прилежащая к нему эктодерма боковой или брюшной области зародыша изменит направление развития и даст начало не кожным покровам, а добавочной центральной нервной системе. В результате, у оперированного описанным образом зародыша, помимо осевых органов, развивающихся, как это имеет место в норме, на спинной стороне, образуется совокупность осевых органов на необычном месте. Подопытный зародыш имеет вид как бы двух соединенных вместе зародышей — первичного и вторичного. Из этих опытов Шпеман сделал вывод, что верхняя губа бластопора и развивающийся из нее хордомезодермальный зачаток обладают способностью воздействовать на прилежащую эктодерму, не только в эксперименте, но и в норме, побуждая ее к превращению в мозговую ткань. Шпеман назвал верхнюю губу бластопора организатором, или организационным центром, а ее формообразовательное воздействие на эктодерму — эмбриональной индукцией (см. также главу 26). За открытие этого явления Шпеман был удостоен в 1935 г. Нобелевской премии.
Позднее было выяснено, что явления индукции свойственны не только амфибиям, но и высшим позвоночным. В частности, было показано, что вторичные осевые органы могут быть индуцированы у куриных зародышей. Аналогичные явления индукции обнаружили и у зародышей кролика.

Ганс Шпеман. 1869–1941.
По представлениям Шпемана и его последователей, все эмбриональное развитие следует рассматривать как цепь индукционных процессов. Верхняя же губа бластопора выступает в роли первичного организатора. Не следует, впрочем, думать, что этот организатор возникает только тогда, когда во время гаструляции образуется спинная губа; он существует у зародыша значительно раньше, вероятно, в еще не начавшем дробиться оплодотворенном яйце в виде определенного участка яйцевой протоплазмы называемого у амфибий «серым серпом». Наложением волосяной лигатуры можно отделить друг от друга два бластомера. Результаты такой изоляции оказываются не всегда одинаковы. Если при первом дроблении борозда, разделяющая яйцо на два бластомера, проходит порез середину «серого серпа», т. е. разделяет яйцевой материал на части, соответствующие правой и левой половинам зародыша, то из каждого бластомера при их изоляции разовьется целый, нормально сформированный зародыш. Если же положение борозды первого дробления таково, что она проходит на границе между будущими спинной и брюшной половинами, то результат изоляции бластомеров будет иным: из одного, соответствующего спинной стороне зародыша, образуется целый организм, а из другого — бесформенное скопление клеток, которое дальше не развивается. Следовательно, в первом случае материал организатора распределяется между бластомерами поровну, а во втором он оказывается только в одном из изолированных бластомеров.
Первичный организатор характеризуется региональностью: отдельные его участки по длинной оси зародыша индуцируют различные структуры. Участок верхней губы бластопора на стадии ранней гаструлы, т. е. будущая передняя часть крыши первичной кишки, индуцирует преимущественно головные структуры, а такой же участок губы бластопора на стадии поздней гаструлы, т. е. будущая задняя часть крыши первичной кишки, вызывает образование туловищных и хвостовых структур.
Из этих опытов первоначально напрашивался вывод, что реагирующая на индукционное воздействие эктодерма совершенно пассивна. Позднее было установлено, что способность эктодермы реагировать на действие индуцирующего агента (эту способность называют компетенцией) изменяется во времени и, подобно организатору, обнаруживает региональность. Вместе с тем экспериментальные данные показывают, что компетенция ранней эктодермы настолько широка, что можно говорить о ее тотипотентности; из эктодермальных клеток удавалось под влиянием индукции получать нейральные, эктомезенхимные, мезодермальные и даже энтодермальные образования. Возникающая в эксперименте нейрализация эктодермы может быть получена даже без воздействия индуцирующего агента. Это явление «самонейрализации» еще недостаточно проанализировано. Предполагают, что оно вызывается частичным цитолизом эктодермы под влиянием неблагоприятных условий или токсических воздействий.
Морфогенез отдельных органов.
После того, как первичный организатор — хордомезодермальный зачаток — индуцировал в спинной эктодерме образование центральной нервной системы, обнаруживается действие других индукторов — организаторов второго и третьего порядка, которое было раньше всего обнаружено на наиболее подробно изученном морфогенезе глаза позвоночных. На стадиях гаструляции и нейруляции под индукционным действием переднего отдела хордомезодермы в нервной пластинке закладывается глазной материал, который затем обособляется от головного мозга — боковые участки переднего мозгового пузыря разрастаются в стороны, образуя глазные пузыри. Последние растут по направлению к эктодерме и вступают в контакт с ее внутренней поверхностью. Из глубокого слоя этих участков эктодермы образуется хрусталик, а из поверхностного — роговица, при этом первоначально округлый глазной пузырь уплощается, а затем образует двуслойную глазную чашу. На первых порах эмбриологи предполагали, что превращение глазного пузыря в чашу происходит в результате давления, оказываемого растущим хрусталиком. Как уже было сказано выше, дело обстоит иначе: образование хрусталика и роговицы есть следствие индукционного влияния глазной чаши на головную эктодерму.
За первыми работами, анализировавшими взаимодействие частей развивающегося глаза (Г. Шпеман, 1901; У. Льюис, 1904), последовало много работ, итоги которых обобщены, в частности, в книге Г.В. Лопашова и О.Г. Строевой (1963).
Примером вторичной индукции является образование зачатка внутреннего уха — слухового пузырька — и развитие вокруг него хрящевой капсулы. Индуктором слухового пузырька является средний мозг, а индуктором слуховой капсулы — слуховой пузырек. Последний факт был установлен Д.П. Филатовым (1916), который показал, что удаление одного слухового пузырька у зародыша жабы приводит к отсутствию хрящевой капсулы на соответствующей стороне тела, а пересадка слухового пузырька, освобожденного от окружающих его мезенхимных клеток, в другое место головы зародыша влечет за собой образование вокруг пересаженного пузырька хрящевой капсулы из мезенхимы, которая в норме в хрящевую ткань не превращается.
Естественно, что подобного рода факты заставили задуматься над вопросом о природе индукционного воздействия. Прежде всего было установлено, что индуцировать нервную систему из любого участка эктодермы тритона могут не только организаторы, взятые от зародышей того же вида но и от зародышей других видов тритона, а также амфибии другого отряда (бесхвостые) и даже животных другого класса (птицы).
Вторичные организаторы, например, глазная чаша как индуктор хрусталика, или слуховой пузырек как индуктор хрящевой слуховой капсулы также не обладают видовой специфичностью. Далее выяснилось, что в эксперименте индуктор может вызывать развитие не только того органа, который под его влиянием возникает в норме, но и совершенно иного. Б.И. Балинский (1927) показал, что у зародыша тритона пересадка слухового пузырька или носовой плакоды под эктодерму бока в области между местами закладки будущих передней и задней конечностей индуцирует в месте трансплантации образование добавочной, пятой конечности. Кстати, здесь также отсутствует видовая специфичность индуктора, и в опыте с равным успехом может быть использован слуховой пузырек лягушки. Способность к индукции передается от организатора к той части, которая образовалась под его влиянием; уже образовавшаяся закладка нервной системы (медуллярная пластинка) в свою очередь способна индуцировать образование медуллярной пластинки при ее пересадке другому зародышу.
Индуцирующие вещества.
Все эти факты привели Шпемана к мысли, что индукционное действие организатора зависит от каких-то вырабатывающихся в нем веществ. Однако в течение десяти лет, прошедших со времени открытия организационного центра в верхней губе бластопора, подтвердить эту гипотезу не удавалось. Впрочем, в 1931 г. Шпеман и его сотрудники показали, что организатор сохраняет индукционную способность даже после механического разрушения составляющих его клеток и после интенсивного воздействия наркотизирующими веществами. Оставалось только не вполне ясным, были ли клеточные элементы организатора разрушены в этих опытах раздавливанием или наркотиком. Однако уже со следующего года экспериментаторы школы Шпемана начали публиковать работы, в которых описывали индукцию посредством несомненно убитых организаторов. Они нагревали индуцирующий материал сначала до 60°, затем даже до 145°, замораживали его, погружали в спирт, проводили через спирты и ксилол в парафин и затем через ксилол и спирты снова в воду. Несмотря на все эти процедуры, после которых в организаторе не оставалось, конечно, ни одной живой клетки, пересадка этого материала в бластоцель приводила к возникновению добавочных медуллярных пластинок. Убитые организаторы вызывали индукцию нервной ткани не только в эктодерме целого зародыша, но и в изолированных кусочках этого листа.
И. Гольтфретер (1933) показал, что лоскут эктодермы ранней гаструлы, помещенный в физиологический раствор, остается неизменным, если же в такой участок эктодермы завернуть кусочек убитого организатора, в ней образуется медуллярная пластинка. Индукционный эффект в подобном опыте можно получить не только от убитого организатора, т. е. верхней губы бластопора, но и от любой убитой ткани, взятой от зародыша взрослого животного и из человеческого трупа. Впрочем, различные ткани взрослых животных, как показали исследования в лабораториях И. Гольтфретера и С. Тойвонена, обладают качественно неодинаковой индукционной способностью: одни из них дают начало одним, другие другим отделам центральной нервной системы; кроме того, они индуцируют не только нервную ткань (нервную трубку), но и образования, развивающиеся из осевой мезодермы. В результате многолетних исследований Тойвонен (1953) пришел к заключению, что индукции в эмбриональном развитии осуществляются, по меньшей мере, двумя различными веществами: одно из них вызывает развитие нервной ткани, другое — мезодермы (последнее вещество Тойвонен выделил из костного мозга).
Для выяснения способа действия индуцирующих веществ практиковалось отделение компетентной эктодермы от источника индукции при помощи фильтров различной толщины и с различными размерами пор. Первоначально представление о переходе активных веществ из индуктора в реагирующую ткань при отсутствии их непосредственного контакта противопоставлялось взгляду, согласно которому для осуществления индукции необходим контакт, не связанный с переходом индуцирующих веществ.
В последнее время наблюдения при помощи электронного микроскопа обнаружили наличие субмикроскопических тяжей, соединяющих в гаструле нижнюю поверхность спинной эктодермы с крышей первичной кишки, в результате чего был сделан вывод, что «индуцирующее действие оказывают диффундирующие агенты, а для создания оптимальных условий для такого перехода клетки должны находиться в тесном контакте при нормальной инвагинации верхней губы бластопора»[107].
Формообразовательный аппарат.
Большинство современных эмбриологов склоняется к выводу, что для осуществления индукции имеют значение как свойства (компетенция) реагирующей ткани (см. главу 26), так и свойства индуктора, хотя механизм индукционного действия продолжает оставаться неизвестным. Подобную точку зрения в 30-х годах развивал Д.П. Филатов, признававший специфический характер формообразовательных влияний одних частей зародыша на другие. Совокупность непосредственно взаимодействующих частей, участвующих в создании целостного органа или системы, Филатов называл формообразовательным аппаратом. Такими формообразовательными аппаратами являются, например, крыша первичной кишки (хордомезодерма) и спинная эктодерма гаструлы, дающие у позвоночных начало всем осевым органам зародыша; глазная чаша и головная эктодерма с прилежащей мезензимой, образующие все части глазного яблока, сетчатку, пигментный эпителий и радужину, белковую оболочку, роговицу и хрусталик.
По мнению Филатова, закономерности развития зародыша могут быть установлены только при таких исследованиях, в которых применяются и аналитический эксперимент, и сопоставление сходных процессов у животных различных систематических групп при учете исторического (филогенетического) развития способов эмбриогенеза.
Регуляционная теория индукции.
Оригинальная трактовка явлений эмбриональной индукции была предложена М.А. Воронцовой (1960) и независимо от нее Б.П. Токиным (1959).
Воронцова рассматривала процессы индивидуального развития как проявление общей всем живым существам способности к регуляции. В случае индукции хрусталика глазной чашей или хрящевой капсулы слуховым пузырьком зачаток (индуктор) дополняется за счет индуцированного им образования до целого органа зрения или слуха. При индукции центральной нервной системы хордомезодермальным зачатком также происходит регулятивное дополнение части до целого и образуется единая система осевых органов, а подчас даже почти полностью сформированный вторичный зародыш.
Сопоставляя результаты индукции, варьирующие от появления слабо дифференцированной нервной ткани до сформирования почти целого добавочного зародыша, Токин рассматривает индукцию как следствие стимуляции индуктором способности соматических клеток к развитию целого организма. Эта способность к «соматическому эмбриогенезу», по терминологии Токина, в норме не реализуется. Ее проявлению препятствуют структурные и функциональные корреляции частей зародыша. Индуктор нарушает эти корреляции, почему следует говорить не об организующем, а о дезорганизующем, или точнее, дезинтегрирующем его действии. Освобождение части зародыша от интегрирующего действия целого приводит к развитию этой части в направлении самостоятельного целого.
* * *
В 50-60-е годы развитие эмбриологии как науки, изучающей закономерности морфогенеза, продолжалось. Однако в связи с общей ориентацией биологических исследований в сторону познания интимных механизмов жизненных явлений, протекающих на молекулярном уровне, внимание эмбриологов все более сосредоточивалось на тех молекулярных процессах, которые лежат в основе реализации наследственной информации в виде свойств и признаков развивающегося и сформированного организма. В результате синтеза ряда дисциплин сложилось новое комплексное направление — биология развития, которое стало ключевым в раскрытии закономерностей индивидуального развития. Рассмотрению проблематики этого направления посвящена глава 26.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК