Глава 10 Цитология
Клетка была открыта во второй половине XVII в. Особенно интенсивно изучение клетки развернулось во второй половине XIX в. в связи с созданием клеточной теории. Клеточный уровень исследования сделался ведущим принципом важнейших биологических дисциплин. Уже в 1858 г. вышла в свет «Клеточная патология» Р. Вирхова, а в 1895 г, — «Клеточная физиология» М. Ферворна. В последней четверти XIX в. изучению клетки посвятила себя целая плеяда первоклассных исследователей — Г. Фоль, О. Бючли, О. Гертвиг, Э. ван Бенеден, В. Флемминг, Э. Страсбургер, Ж. Карнуа и многие другие. Благодаря их работам было детально изучено строение клеточного ядра, проведен цитологический анализ таких важнейших биологических процессов, как митоз, мейоз, оплодотворение. Однако цитологии как самостоятельной научной дисциплины еще не существовало, поскольку во всех этих исследованиях клетка рассматривалась только в качестве составного элемента организма.
Однако именно к концу XIX в. относится пробуждение специального интереса к клетке как самостоятельной единице, имеющей общебиологическое значение, так как, с одной стороны, обнаружилось, что клетки обладают рядом общих свойств независимо от их происхождения, а с другой стороны, выяснилось, что разные клетки в зависимости от выполняемой ими функции имеют неодинаковые строение и свойства. К углубленному изучению клетки, ее внутренних структур побуждали, в частности, интересы зарождавшейся генетики, искавшей конкретный материальный субстрат наследственности.
Было установлено, что клетки являются не только структурными элементами многоклеточного организма, но могут существовать и самостоятельно. Яйцо, дающее начало всем многообразным структурам будущего организма, представляет собой клетку. Простейшие, несмотря на разнообразие и подчас сложность строения — также отдельные клетки. Эти факты привели к представлению, что клетка — это элементарный организм, живущий самостоятельно, или этап развития будущего многоклеточного организма. Возникала необходимость всестороннего изучения этой универсальной жизненной единицы.
К концу XIX и началу XX в. относится появление первых больших сводок о клетке и ее роли в основных жизненных процессах. Это капитальный труд И. Деляжа «Структура протоплазмы, теории наследственности и важнейшие проблемы общей биологии» (1895), книга Э. Вильсона «Клетка и ее роль в развитии и наследственности» (1896), содержащая богатый цитологический материал, а также монографии А.Г. Гурвича «Морфология и биология клетки» (1904) и М. Гейденгайна «Плазма и клетка» (1907). Основным содержанием монографии Гурвича является освещение важнейших свойств клетки и ее жизнедеятельности. В соответствии с этим основные разделы этого издания названы «Статика и динамика клетки», «Метаболизм клетки», «Размножение клетки» и «Клетка как организм и индивидуум». В книге Гейденгайна особое внимание уделено протоплазме. Она рассматривается как важнейший субстрат, из которого построены клетки и который определяет их основные жизненные свойства.
Программу цитологических исследований XX в. четко сформулировал С.Г. Навашин, утверждая, что «задачей нового столетия является выяснить и точнее разграничить свойства клетки»[88].
Новые методы и средства исследования.
В XIX в. в основном изучали мертвую клетку после ее фиксации и окраски. При этом удавалось описать ряд ее органоидов и структур — митохондрии, аппарат Гольджи, клеточный центр и т. д., но более точное знание структуры и особенно функции клетки при таком методе исследования было недостижимо; кроме того, отсутствовала уверенность, что наблюдаемые на препарате картины соответствуют прижизненным.
Это очень отчетливо проявилось, например, во время спора, возникшего в конце XIX в., о тонкой структуре цитоплазмы, когда были предложены различные теории строения протоплазмы — зернистого (Р. Альтман), фибриллярного (В. Флемминг) и пенистого, или ячеистого (О. Бючли). А. Фишер (1899) обнаружил беспредметность этого спора. На белковых моделях он показал, что в зависимости от условий опыта возможно получение картин, соответствующих всем трем теориям строения цитоплазмы.
Возникла необходимость, не отказываясь от этого пути исследования, совмещать его с наблюдением живой клетки. Систематическое изучение живых клеток началось только в начале XX в. благодаря открытию возможности переживания клеток в культуре вне организма.
Метод эксплантации был впервые применен американцем Р. Гаррисоном в 1907 г. На кусочках зачатка нервной системы лягушки, помещенных в каплю лимфы, он наблюдал переживание Клеток в течение некоторого времени и даже появление и рост у них нервных отростков. Метод культуры ткани был затем значительно усовершенствован в работах А. Карреля (1911–1915), М. Берроуса (1910, 1912), А.А. Максимова (1916, 1925), В. и М. Льюисов (1911 и позднее), А. Фишера (1925), А.В. Румянцева (1932) и многих других.
Широкое и полноценное использование культивируемых вне организма клеток для цитологических исследований стало возможным благодаря введению в 50-х годах метода клеточных культур на жидких питательных средах.

Росс Гренвил Гаррисон. 1870–1959.
Клеточные культуры представляют собой популяции самостоятельных клеток, которые только вторично и далеко не всегда устанавливают между собой протоплазматические связи, однако оказывают взаимное гуморальное влияние. Клетки однослойных культур служат превосходным объектом и для проведения микроскопических исследований с использованием замедленной (цейтрафферной) микрокиносъемки. С помощью микрокиносъемки изучены движение, деление, фагоцитоз и многие другие нормальные и патологические процессы в клетках.
Наряду с применением клеточных культур в цитологии, особенно в последние годы, широко используются культуры органов, клетки которых входят в состав целых эмбриональных зачатков или фрагментов органов взрослых животных (А. Тома, 1970).
Существенное значение для развития метода прижизненного изучения клетки имело усовершенствование оптических средств исследования. К их числу относится прежде всего темнопольная микроскопия («ультрамикроскопия»), практическое применение которой стало возможно после изобретения в 1910 г. Г. Зидентопфом кардиоид-конденсора. Важные цитологические наблюдения с использованием темнопольной методики сделал Н.М. Гайдуков (1916).
В 40-50-х годах широкое распространение получила фазово-контрастная микроскопия. Ее разновидностью явилось аноптральное устройство, схему которого разработали А. Вильска (1953) и М.А. Пешков.
Прижизненному изучению клетки содействовало также развитие люминесцентной (флуоресцентной) микроскопии. В основе этого метода лежит открытие А. Келером (1904) флуоресценции объектов в темном поле при освещении их ультрафиолетовыми лучами (УФ). Широкое применение в цитологии люминесцентного микроскопа, сконструированного Келером и Зидентопфом (1908), стало возможным после того, как М. Гайтингер разработал метод окраски биологических объектов флуорохромами — веществами, обладающими способностью светиться после их УФ-облучения. С помощью люминесцентной микроскопии получены ценные сведения о строении, обмене веществ и функционировании нормальных и патологически измененных клеток (раковых, после воздействия вирусов и т. д.).

Уоррен Льюис. 1870–1964.
Одно из наиболее важных направлений флуоресцентной микроскопии связано с применением метода флуоресцирующих антител, впервые предложенного в 1941–1942 гг. А. Кунсом и сотрудниками (1956) для выявления чужеродного антигена. Содержащиеся в сыворотке антитела связываются с флуорохромами. Меченные таким образом антитела соединяются с соответствующими антигенами и позволяют определить локализацию как бактериальных и вирусных антигенов, так и различных белков, вызывающих образование соответствующих антител. Метод флуоресцирующих антител позволил изучить локализацию и динамику накопления специфических белков, в частности ряда ферментов, появление белковых продуктов эмбриональной дифференцировки, топографию вируса и динамику его накопления в клетке, реакцию иммунокомпетентных клеток на проникновение в организм инфекции и т. д.
Основным методом изучения клетки в прижизненном состоянии стала микрургия, дающая возможность производить на клетках разнообразные операции. Разработка достаточно совершенной техники операций на клетках относится к началу XX в., когда был сконструирован микроманипулятор. Этот инструмент позволил извлекать из клетки отдельные органоиды (в частности, хромосомы), измерять электрические потенциалы с помощью микроэлектродов, вводить в клетку разнообразные вещества, бактерии, ядра и другие компоненты сходных или чужеродных клеток. Микроманипулятор был создан в самом начале XX в. одновременно С. Схоутеном (1901) в Нидерландах, М. Мак-Клендоном и М. Барбером (1901) в США. С.С. Чахотин (1959) использовал изобретенный им микрооператор (1912) для строго локализованного воздействия на клетку пучка УФ (лучевой микроукол). С помощью локального воздействия УФ удавалось нарушить двойное лучепреломление в ахроматическом веретене (клетки подсолнечника), повредить участки митотического аппарата (в клетках тритона) и т. д. Основные работы с микроманипулятором были выполнены во втором десятилетии XX в. американским исследователем Р. Чемберсом.
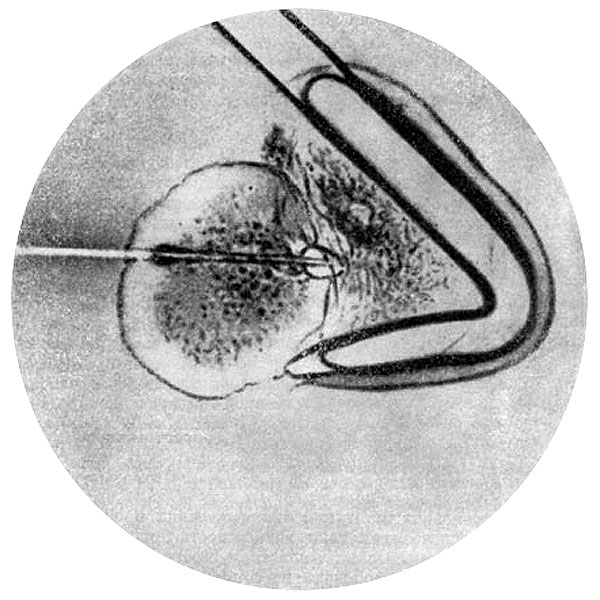
Извлечение ядра из амебы с помощью микроинструмента, по П. Фонбрюну. 1951.
Наряду со значительными успехами в изучении живой клетки XX в. ознаменовался крупными достижениями и в исследовании клетки фиксированной. Первостепенную роль здесь сыграли методы цитохимии (гистохимии).
Основной задачей цитохимии является выяснение химической природы клеточных структур и продуктов жизнедеятельности клетки, а также расшифровка происходящих в ней биохимических процессов. Цитохимические методы исследования дают возможность топографически изучить локализацию ряда химических веществ и процессов. Применение всех цитохимических реакций основано на природной или искусственно создаваемой окраске изучаемых веществ. Примером цитохимических реакций первого рода является изучение пигментов в клетке, а применение реакций второго рода основано на избирательной окраске некоторых внутриклеточных веществ (например, окраска нейтрального жира Суданом) или на выпадении в результате химической реакции нерастворимого осадка какого-либо вещества. Так, о наличии и локализации активности кислой фосфатазы судят по локализации осадка сернистого свинца в присутствии субстрата ферментативной реакции (?-глицерофосфата).

Роберт Чемберс. 1881–1957.
Гистохимия и цитохимия берут свое начало от исследований француза Ф. Распайля, выполненных еще в 30-х годах XIX в. Он, по-видимому, первым использовал йодную реакцию на крахмал для его обнаружения в растительной клетке. Интенсивное развитие цитохимии началось в 30-40-х годах XX в., когда были предложены методы обнаружения важнейших химических компонентов клетки и опубликован ряд соответствующих руководств и статей. Основополагающее значение имели труды Л. Лизона (1936, 1953), а также работы К. Бенсли (1941), Д. Пайка (1950), Дж. Гомори (1952), Э. Пирса (1962) и многих других. Современные цитохимические методы дают возможность изучать биохимические процессы не только в неразрушенной, но иногда и в живой клетке. С помощью этих методов оказывается возможным связать изучаемый биохимический процесс с определенной клеточной структурой и прослеживать его в динамике.
Одной из важнейших цитохимических реакций, введенных в первой половине XX в., является открытая Р. Фельгеном (1924) специфическая реакция на ДНК. В 1940 г. Ж. Брате предложил специфическую окраску РНК пиронином и метиловым зеленым. В последующие годы были разработаны методики для обнаружения аминокислот, белков, углеводов, витаминов и определения активности окислительных, гликолитических и других ферментов.
Наряду с цитохимическими методами использовалось также дифференциальное центрифугирование, дающее возможность изолировать из клетки и разделить входящие в ее состав органоиды. Впервые дифференциальное центрифугирование было применено в 30-х годах К. Бенсли и Н. Херром для выделения и обособленного анализа митохондрий.
Существенную роль в изучении обмена веществ клетки сыграл введенный в 1940 г. шведским ученым Т. Касперсоном метод количественного учета веществ (главным образом нуклеиновых кислот) по поглощению в изучаемом объекте УФ-лучей определенной длины волны. Этот точный метод дает возможность получить сведения о количестве и распределении нуклеиновых компонентов клетки и некоторых других звеньев ее метаболизма.
При изучении динамики обмена веществ клетки большое распространение получило использование радиоактивных изотопов и радиоавтографии (Г. Хевеши, 1950; Дж. Бойд, 1957). Оно основано на том, что радиоактивные изотопы (Н3, С14, S36, N15), подведенные к клеткам в процессе метаболизма, включаются в состав некоторых важных химических компонентов (ДНК, РНК и белков). Если покрыть гистологический срез или пластинку, на которой растет однослойная клеточная культура, жидкой фотоэмульсией, то вещества, излучающие главным образом частицы, вызовут восстановление бромистого серебра. По выявившимся зернам металлического серебра можно получить представление о локализации и количестве изучаемого вещества (Л.Н. Жинкин, 1959). Оказалось, что использование радиоактивных изотопов дает возможность изучить динамику синтеза ДНК (М. Говард, С. Пелк, 1951). Особенно большое значение для цитологии радиоавтография приобрела после того, как в 1959 г. Г. Квастлером и Ф. Шерманом был разработан метод радиоавтографии с использованием предшественника ДНК — Н3-тимидина.
Наконец, очень важным методическим приемом, коренным образом изменившим направление цитологии, явилась электронная микроскопия. Проектирование и создание электронного микроскопа относятся к 30-м годам и связано с именами преимущественно немецких физиков (Ф. Вольф, М. Кнолль, Э. Руска, Э. Брюхе, Г. Иохансон). Этот метод в сочетании с использованием ультратонких срезов позволяет изучить ультрамикроскопическую структуру всех основных органоидов (современные электронные микроскопы обладают разрешающей способностью в несколько микронов). Применение электронного микроскопа привело к перестройке классических представлений не только о строении клетки, но и о ходе многих процессов ее жизнедеятельности.
Наибольшие результаты дает сочетание высокой техники электронномикроскопического исследования с комбинированным применением радио- автографических и цитохимических методов. Используя связывание тяжелых металлов (свинца, кадмия) определенными компонентами клетки, можно выяснить ультрамикроскопическую локализацию ферментативной активности (сукцинатдегидрогеназной, фосфатазной и т. д.) Точно так же, определив на ультратонких срезах место локализации металлического серебра после предварительного введения радиоизотопов, можно получить данные об участках метаболической активности клетки.
В настоящее время в области ультраструктуры клетки после работ А. Клода (1940), Г. Палада (1952, 1953), Ф. Шёстранда (1956), К. Портера (1961) и других достаточно отчетливо выяснены как общая схема строения клетки, так и организация ее отдельных структурных элементов.
Сравнительно недавно были обнаружены, описаны и идентифицированы эндоплазматическая сеть, мембранная система, лизосомы, фагосомы, микротельца, микротрубочки и другие структуры. Ядро, ядрышко, митохондрии, комплекс Гольджи, клеточный центр, плазматическая и ядерная оболочки, известные ранее, описаны при помощи новых методических приемов во всех ультраструктурных деталях. С помощью электронной микроскопии удалось выяснить морфологический субстрат основных процессов, происходящих в клетке, — пассивного и активного проникновения в нее различных веществ, слияния и рекомбинации мембран, выделения секретов, фагоцитоза и пиноцитоза, движения и сократимости, проведения нервных импульсов (см. главу 11).
Таким образом, развитие техники цитологических исследований и введение некоторых принципиально новых методических приемов одновременно послужило предпосылкой развития цитологии и предопределило формирование основных направлений, по которым пошло ее дальнейшее развитие.
Физико-химическое изучение клетки.
Одним из основных направлений цитологии в первой трети XX в. было изучение физико-химических свойств клетки. Значительные успехи были достигнуты в исследовании физических свойств и коллоидного состояния протоплазмы. Были получены исчерпывающие данные о ее основных физических свойствах — вязкости, эластичности, электрическом заряде, поверхностном натяжении, концентрации водородных ионов, проницаемости, чувствительности к различным (в частности, лучевым) воздействиям и т. д. Работы Р. Чемберса, Т. Петерфи, Ж. Лёба, В. Зейфрица, Ф. Вебера и многих других заложили основы этой области цитологии. Результаты исследований получили отражение в крупных сводках и руководствах. Таковы: «Физическая химия клетки и тканей» (1911) Р. Гёбера, «Коллоидная химия протоплазмы» (1929) Л. Гейлбруна, «Проблема проницаемости» (1929) Е. Гельхорна, «Протоплазма» (1936) В. Зейфрица и многие другие.
Физико-химическое изучение клетки получило широкое развитие в 20-х годах и в Советском Союзе. Центром исследований в этой области стал московский Институт экспериментальной биологии, руководимый Н.К. Кольцовым. Именно здесь были выполнены важные работы самого Н.К. Кольцова, С.Н. Скадовского, Г.В. Эпштейна. В итоге было детально изучено влияние на клетку водородных ионов, заложено представление о физико-химических основах раздражимости пигментных, мускульных и железистых клеток. Особенно много внимания Кольцов уделил физико-химическому анализу формы клеток. Именно под этим углом зрения выполнены его классические исследования строения спермиев десятиногих раков, сократительного стебелька сувойки и др. Основной задачей Кольцов ставил изучение структур и элементарных процессов, обеспечивающих всю сложность жизни клетки. Книга Кольцова «Организация клетки» (1936), обобщившая результаты его 30-летней работы и отразившая его теоретические представления, внесла существенный вклад в цитологию. Основным выводом из исследований Кольцова и его школы явилось представление о твердом скелете, который в той или иной форме присутствует в любой клетке и определяет ее организацию.
Физико-химическое изучение клетки велось также в Институте биохимии им. А.Н. Баха (Д.Л. Рубинштейн, В.А. Дорфман и др.). Выполненные здесь исследования касались вопросов проницаемости клетки, электрометрии (в первую очередь изучения клеточных потенциалов), а также ее диэлектрических свойств.
Для выяснения физико-химических свойств клетки много сделал в Московском университете А.В. Румянцев, который справедливо может считаться одним из основателей московской цитологической школы. Работы Румянцева, начало которых относится к первой половине 20-х годов, посвящены изучению строения и свойств живой клетки при воздействии разнообразных химических и физических факторов. Они содержали значительную долю здорового скептицизма по отношению к результатам, полученным на фиксированных и окрашенных препаратах. И действительно, некоторые классические клеточные структуры оказались всего лишь артефактами. Так, уже в первой большой цитологической работе (1925), посвященной изучению протоплазмы корненожек, Румянцев выяснил природу так называемых хромидий — цитоплазматических образований, окрашивающихся подобно хроматину, которые одно время считались важными органоидами клетки. Румянцев показал, что речь идет об остатках разрушенных митохондрий и аппарата Гольджи, вряд ли имеющих какое-либо физиологическое значение.
Метод культуры ткани Румянцев использовал для решения не гистологических или физиологических, а именно цитологических задач. Данные, полученные на клетках культуры ткани, отражены в серии работ, вышедших в 1927–1929 гг. и объединенных общим названием «Цитологические исследования в тканевых культурах». Много цитологических данных содержится в монографии Румянцева «Культуры тканей вне организма и их значение в биологии» (1932).
На клетках культуры ткани были исследованы аппарат Гольджи, митохондрии, структуры ядра, строение цитоплазмы в норме и при воздействии разнообразных физических и химических факторов — изменении реакции среды, воздействии некоторых ионов и солей. Несмотря на кажущуюся пестроту вопросов, эти исследования имели свою внутреннюю логику — изучалась протоплазма как субстрат жизненных явлений, выявлялись пределы ее реактивности, устанавливались причины ошибок, основанных на некритическом отношении к результатам изучения статических картин.
Некоторые исследователи уже в первые десятилетия XX в. пытались связать данные о физико-химических свойствах клетки с ее функциями. Новое физиологическое направление в цитологии особенно отчетливо выявилось в работах Д.Н. Насонова. В начале 20-х годов он приступил к изучению одного из труднейших для истолкования органоидов клетки — аппарата Гольджи. Открытая в конце XIX в., эта структура привлекала к себе пристальное внимание исследователей, однако вопрос о реальном существовании и особенно функциональном значении аппарата Гольджи долгое время оставался открытым. В результате исследования, проведенного на различных железах амфибий, Насонов (1923, 1963) пришел к заключению, что аппарат Гольджи является основным органоидом внутриклеточной секреции. В своих исследованиях он использовал экспериментальный подход — животным вводился пилокарпин, стимулирующий клеточную секрецию, что приводило к освобождению клетки от секрета и позволяло исследовать роль аппарата Гольджи в возникновении процесса секреции.
Почти одновременно с изучением аппарата Гольджи-Насоновым американский исследователь Р. Боуэн (1924, 1925) выполнил серию работ, посвященных сперматогенезу у насекомых и секреторному процессу в железах моллюсков, амфибий, птиц и млекопитающих. Он получил результаты, полностью совпавшие с выводами Насонова: аппарат Гольджи продуцирует клеточный секрет. Одновременно Боуэн показал существование различий в структуре этого органоида у беспозвоночных и позвоночных и высказал предположение, что такие различия связаны с особенностями секреции этих групп.
В одной из работ Насонова (1926) было исследовано накопление кислого витального красителя (трипанового синего) в клетках печени и почек различных позвоночных. При этом было показано, что краситель концентрируется в области аппарата Гольджи, не вызывая каких-либо его морфологических изменений. Полученные результаты дали автору основание считать, что деятельность аппарата Гольджи сводится к избирательной концентрации находящихся в клетке веществ, независимо от того, образуются ли они на месте или поступают в клетку извне. Эти представления Насонова о функции аппарата Гольджи в основном сохраняют свое значение и в настоящее время.
Большое число исследований первой трети XX в. было посвящено витальному окрашиванию. Биологическое значение этого метода заключается в том, что он дает возможность прижизненно изучить закономерности поступления, накопления и выделения входящих в клетку веществ. Иными словами, он позволяет создать модель процессов обмена веществ. Витальное окрашивание (его феноменология и механизм) детально описаны в работах Г. Эванса и В. Шулемана (1915) и В. Меллендорфа (1920). Большую роль в развитии этого направления исследований сыграли работы Н.Г. Хлопина (1927), выдвинувшего представление о криномах — структурах, предсуществующих в клетке или возникающих заново, которые связывают попадающие в клетку вещества, в частности красители. В дальнейшем ряд важных исследований по изучению реакции клетки на введение анилиновых красителей был выполнен Б.В. Кедровским (1936), разработавшим представление о роли гранул как сегрегационного аппарата клетки. С развитием электронно-микроскопических исследований выяснилось, что гранулярный аппарат клетки тесно связан с ультрамикроскопическими структурами — лизосомами, которым в настоящее время отводится важная роль в физиологии нормальной и патологически измененной клетки.

Дмитрий Николаевич Насонов. 1895–1957.
Значение витальной окраски для характеристики физиологического состояния клетки особенно четко показано в работах Д.Н. Насонова и В.Я. Александрова, сформулировавших теорию паранекроза (1940). Она основывалась на следующих наблюдениях: в норме основные красители откладываются в виде гранул в цитоплазме, ядро остается неокрашенным и интенсивность гранулообразования сравнительно невелика. При умеренном раздражении, связанном с уменьшением дисперсности белков цитоплазмы и с повышением ее сорбционных свойств, количество и размеры гранул возрастают. Значительное повреждение клетки сопровождается потерей способности к гранулообразованию, появлением диффузной окраски цитоплазмы и прокрашиванием структур ядра. Эти изменения, наблюдаемые при разнообразных внешних воздействиях, являются монотонным ответом на них любой клетки и соответствуют ее своеобразному некробиотическому состоянию, которое было названо авторами паранекрозом. Ранние стадии этого состояния обратимы. В гибнущей клетке вследствие денатурации белков диффузная окраска цитоплазмы и ядра сохраняется.
Авторы рассматривают образование гранул (или оседание красителя на предсуществующих структурах) как проявление действия защитного механизма, с помощью которого из основной цитоплазмы удаляются посторонние или вредные вещества. В случаях, когда клетка повреждена, этот защитный механизм нарушается и гранулярная окраска становится уже невозможной. Таким образом, гранулообразование может служить одним из достаточно тонких индикаторов физиологического состояния клетки не только при естественных колебаниях ее жизнедеятельности, но и в тех случаях, когда на клетку действуют извне разнообразные (в том числе повреждающие) факторы.
Теория паранекроза дает возможность объяснить однообразие ответа клетки на разнообразные внешние воздействия. Она послужила основой при изучении другого, очень важного раздела физиологии клетки — проблемы проницаемости.
Изучение проницаемости.
Эта проблема начиная с 20-х годов XX в. привлекла к себе большое внимание цитологов и породила множество гипотез, толкований и споров.
Представления Э. Пфеффера и Г. де Фриза (конец XIX в.) положили начало теории клеточной проницаемости, в которой обменные процессы, протекающие в клетках, ставились в тесную связь с наличием полупроницаемой мембраны. В ее защиту выступил, в частности, немецкий биохимик Е. Овертон (1895, 1899, 1902). Не располагая прямыми доказательствами существования полупроницаемой мембраны, Овертон на основании косвенных физико-химических данных выдвинул липидную (липоидную) теорию клеточной проницаемости, согласно которой плазматическая мембрана представляет собой пленку из жироподобных веществ — липоидов. Постепенно стали накапливаться факты, свидетельствовавшие о несоответствии количеств проникающих в клетку веществ их растворимости в липоидах, а также поступлении в нее веществ, в липоидах совершенно не растворимых. Пытаясь спасти теорию Овертона, его соотечественник А. Натансон (1904) высказал предположение о мозаичном строении мембраны. В 1908–1913 гг. немецкий физиолог В. Руланд развил получившую широкую известность теорию ультрафильтра, или «сита», согласно которой в полупроницаемой мембране имеются поры определенного диаметра, через которые в клетку могут проникать лишь молекулы соответствующих размеров. Однако, как справедливо отметили Р. Коллайдер и Г. Берлюид (1933), теория Руланда не объясняла того факта, что проницаемость многих растительных клеток растет для веществ гомологического ряда по мере увеличения в них числа атомов углерода. Поэтому они предложили соединить липидную теорию с теорией ультрафильтра, допустив тем самым существование двух разнородных механизмов, регулирующих проникновение в клетку молекул различной природы. Эта точка зрения в 30-40-е годы получила поддержку многих биологов (см. главу 11).
Но и в таком, усовершенствованном виде мембранная теория оказалась неспособной объяснить целый ряд явлений и прежде всего основной факт — стационарное распределение веществ в клетке, качественно отличное от состояния простого водного раствора. Слабой стороной этой теории было также игнорирование всех остальных компонентов клетки. В 1907 г. ее впервые подвергли критике М. Фишер и Г. Моор, которые рассматривали поступление в клетку различных веществ не как следствие осмотических и диффузионных закономерностей, а как результат коллоидно-химических процессов, протекающих в самой протоплазме. Сомнения в решающей роли плазматической мембраны в обеспечении проницаемости были высказаны В.В. Лепешкиным (1924, 1930) и Д.А. Сабининым (1920–1925); последний подробно исследовал зависимость проницаемости от реакции среды.
Однако наиболее обоснованные возражения против мембранной теории сформулировали в 30-е годы Д.Н. Насонов и В.Я. Александров, а позднее А.С. Трошин (1956). Они разработали сорбционную теорию проницаемости. Согласно этой теории, решающая роль в распределении веществ, проникающих в клетку, принадлежит сорбционным отношениям, устанавливающимся между протоплазмой клетки в целом и окружающей средой. По удачному выражению А.С. Трошина, сорбционную теорию можно было бы назвать протоплазматической теорией проницаемости. Сорбционная теория основывается на следующих положениях: растворимость веществ в протоплазме должна отличаться от растворимости в обычной воде; важнейшим фактором распределения веществ в клетке является их адсорбция и химическое связывание в протоплазме.
Первоначально казалось, что обе теории совершенно несовместимы. Позже, однако, выяснилось, что их сближение возможно.
Ультраструктура клетки.
Широкое использование электронной микроскопии привело к тому, что было существенно дополнено, а в некоторой степени и изменено, традиционное представление о строении клетки.
В настоящее время ведущим является представление о мембранно-вакуолярной системе и об основном матриксе, в который эта система погружена (Ф. Хагено, 1958). Важнейшие процессы жизнедеятельности клетки очень хорошо согласуются с этой схемой ее субмикроскопического строения.
Для характеристики современного уровня знаний об ультраструктуре клетки весьма показательна эволюция в области представлений об отдельных структурах клетки. С помощью дифференциального центрифугирования первоначально удалось выделить только несколько основных фракций, подлежащих дальнейшему анализу, — ядра, митохондрии и микросомы. В последней фракции были найдены обрывки эндоплазматической сети частично с сидящими на них рибосомами и многочисленные, точнее не идентифицированные, гранулы, получившие сборное название цитоплазматических частиц. В последнее время было выяснено, что в состав этой группы входят совершенно гетерогенные образования, различающиеся как по своей ультраструктуре, так и по функции. Так, были описаны лизосомы (К. Де-Дюв, 1963), носители протеолитических ферментов. Им принадлежит, по-видимому, важная роль аппарата защиты клетки и уничтожения ее поврежденных и отмирающих частей. Характерная особенность лизосом — высокая активность в них кислой фосфатазы и некоторых нуклеаз.
В качестве самостоятельных структур были выделены микротельца, чрезвычайно широко распространенные в клетках животных и, видимо, растений. Они оказались тесно связанными с каталазой, уратоксидазой и оксидазами аминокислот (3. Грубан, М. Рехцигл, 1969). Идентифицировать эти и некоторые другие структурные элементы клетки удалось благодаря сочетанию морфологического и цитохимического методов.
Химическая цитология.
В 30-х годах в цитологии появилось новое направление, которое может быть названо химической цитологией. Его задачей стало изучение биохимических процессов, протекающих в клетке.
Одна из первых цитохимических работ была выполнена Б.В. Кедровским (1934). Применив прижизненную окраску основными красителями, он обнаружил присутствие в клетке базофильных веществ («кислых анаболитов», по его выражению), участвующих в процессах белкового синтеза. Первоначально Кедровский считал «кислые анаболиты» белками, содержащими фосфатные группы. Но уже скоро он пришел к заключению, что это нуклеиновые кислоты, динамика, локализация и огромное биологическое значение которых впоследствии были изучены Ж. Браше, Т. Касперссоном и рядом других иностранных и советских исследователей.
В течение сравнительно короткого времени благодаря использованию цитохимических (в частности, цитоспектрофотометрических) методов было накоплено огромное количество фактов, свидетельствующих о локализации ДНК в ядрах, а РНК — в цитоплазме всех исследованных клеток — от бактерий и простейших до человека.
Для цитохимии углеводного обмена большое значение имели начатые еще в 30-х годах работы А.Л. Шабадаша (1937, 1948), выполненные на клетках нервной системы, крови, клеточных культурах и т. д. Шабадаш предложил очень удачный метод гистохимического обнаружения гликогена путем его окраски основным красителем — фуксином, после окисления этого углевода периодатами (ШИК-реакция; в зарубежной литературе этот метод именуется ПАС-реакцией). Гистохимическое обнаружение гликогена позволило выявить динамику этого вещества даже при незначительных функциональных изменениях в нервной клетке, в ряде клеток на первых этапах лучевого поражения, в процессе регенерации, при вирусной инфекции в клетках культуры ткани и т. д. Так как гликоген является важным не только энергетическим, но и пластическим компонентом клеточного обмена, определение количества и формы его отложения (десмо- и лиогликоген, по Шабадашу) является одним из наиболее чувствительных критериев физиологического состояния клетки.
Применение дифференциального центрифугирования дало возможность, с одной стороны, выяснить химический состав определенных структурных компонентов клетки, с другой — связать химические вещества, входящие в ее состав, с определенными органоидами. Начиная с 50-х годов интересы переместились в область изучения происходящих в клетке биохимических процессов, в, частности биосинтеза. Раскрытие происходящих в клетке биосинтетических процессов связано с возникновением и развитием молекулярной биологии (см. главу 23). Были предприняты также попытки построить на основе отдельных фактов обобщенное представление о физиологии и биохимии клетки (например, Б.В. Кедровский, 1959).
Новое направление исследований позволило путем использования современных методов микроскопии связать все этапы биосинтеза с определенными структурами клетки. Так, было выяснено, что синтез рибосомной РНК происходит в ядрышке, которое можно рассматривать как участок, в котором берут начало основные синтетические процессы клетки. Мысль о роли ядрышка в синтезе специфического белка была впервые высказана Т. Касперссоном (1950), а впоследствии подтверждена рядом исследователей. Всесторонний анализ ядрышка и его функции содержится в монографии Г. Буша и К. Сметаны (1970). С рибосомами связан центральный этап этих процессов — синтез специфических белков. Сами рибосомы были впервые описаны Г. Паладом (1953) под названием плотных частиц или гранул. Несколько позже их удалось выделить из клетки и определить содержание в них РНК (Г. Палад, П. Сикевиц, 1956).
Новые исследования подтвердили и уточнили одно из основных положений классической цитологии о существовании тесных взаимоотношений между ядром и цитоплазмой. Обнаруженные биохимическими методами связи получили подтверждение в морфологических данных. Электронная микроскопия показала, что в ядерной оболочке имеются поры, благодаря которым ядро оказывается в прямой связи с эндоплазматической сетью, т. е. с вакуолярной системой всей клетки; данные цитохимического анализа позволяют определить, какие именно вещества, как и когда поступают из ядра в цитоплазму и, вероятно, при определенных условиях, в обратном направлении.
Результаты цитохимического и ультраструктурного анализа клетки выявили наличие ряда точек соприкосновения между, казалось бы, антагонистическими мембранной и сорбционной теориями проницаемости. Современные исследования роли мембран как важнейшего структурного элемента клетки, показывают, что вещества, проникшие через плазматическую оболочку, встречаются внутри клетки с рядом других мембран, через которые они должны будут проникнуть раньше, чем достигнут протоплазматических структур. В то же время сорбционные способности различных органоидов клетки, несомненно, различны, и это еще больше подтверждает значение сорбционного эффекта в обеспечении клеточной проницаемости.
Энергетика клетки.
В настоящее время достаточно подробно изучена энергетика клетки. Она оказалась теснейшим образом связанной со структурой и функцией митохондрий.
Митохондрии широко изучались цитологами, особенно в конце XIX и начале XX в. Было предложено несколько весьма совершенных методов их окрашивания, в том числе суправитальное окрашивание янусом зеленым (Л. Михаэлис, 1900). Была установлена их чрезвычайно высокая лабильность и подверженность влиянию ряда физиологических и патологических факторов. Тем не менее, их функции оставались неясными. О. Варбург (1913; Нобелевская премия, 1934) выдвинул гипотезу о связи дыхания с клеточными гранулами, однако эта гипотеза не нашла отклика у цитологов. Д. Кейлин и Э. Хартри (1940) показали, что перенос электронов осуществляется цитохромами, содержащимися во внутриклеточных частицах.
Интенсивное изучение митохондрий в 40-50-х годах связано с работами А. Клода (1945, 1946), Г. Хагебума (1948), В. Шнейдера (1948), Д. Грина (1965), А. Ленинджера (1966) и др. Ультраструктура митохондрий была детально исследована Г. Паладом (1952). В 1949 г. Э. Кеннеди и А. Ленинджер выяснили, что в митохондриях происходит окислительное фосфорилирование. Эти органоиды катализируют трансформацию энергии и служат местом локализации окислительных ферментов. Вместе с тем было установлено, что они обладают осмотической, сократительной, регуляторной и генетической функциями. Клод удачно назвал митохондрии «силовыми станциями» клетки.
Современные исследования по энергетике клетки установили прогрессирующую дифференциацию структур и функций. Как уже было сказано выше, многие компоненты суммарной фракции микросом имеют прямое отношение к биохимическим, в частности окислительным процессам в клетке. Выяснилось, например, что микротельца (иногда их называют пероксисомами) являются носителями ряда важных окислительных ферментов. Дальнейший ультраструктурный анализ митохондрий показал, что эти органоиды имеют сложную внутреннюю структуру: поперечные выросты (кристы) разделяют митохондрию на отдельные внутренние камеры, почему в ней одновременно могут происходить различные, иногда даже антагонистические, процессы. В то же время биохимические и молекулярно-биологические исследования приводят к заключению, что из всех многочисленных функций митохондрий основной является контроль и интеграция отдельных процессов (особенно ферментативных). Митохондрия представляет собой не только «силовую станцию» клетки, но и важнейший органоид регулировки энергетических процессов. Имеются данные, свидетельствующие о том, что митохондрии, быть может, являются одним из факторов нехромосомной передачи наследственных свойств (Т.Б. Казакова, 1965).
Проблема клеточного деления.
К концу XIX в. цитологи располагали почти исчерпывающими знаниями о морфологической стороне митоза. Дальнейшее пополнение данных о клеточном делении шло главным образом за счет изучения наиболее примитивных организмов. Был детально изучен процесс деления у прокариотных (не имеющих оформленного ядра) организмов (бактерии), генетически близкий к митозу (М.А. Пешков, 1966), а также митоз у простейших (И.Б. Райков, 1967), где были найдены крайне своеобразные формы этого процесса. У высших организмов морфологическое изучение митоза шло в основном по линии исследования этого процесса в динамике на живых объектах с помощью микрокиносъемки. В этом отношении большое значение имели работы А. Байера и Дж. Моле-Байер (1956, 1961), выполненные на клетках эндосперма некоторых растений.
Однако подавляющее большинство работ XX в. касалось физиологии клеточного деления, и именно в этом разделе проблемы были достигнуты наибольшие успехи. В сущности, неизученным оставался вопрос о причинах и контролирующих факторах митоза. Основоположником этого направления исследований был А.Г. Гурвич.
Уже в монографии «Морфология и биология клетки» (1904) Гурвич высказал мысль, что должны существовать факторы, обусловливающие возникновение митоза, причем они скорее всего связаны с состоянием самой приступающей к делению клетки. Эти пока еще очень общие представления получили развитие в серии дальнейших исследований Гурвича, обобщенных в монографии «Проблема клеточного деления с физиологической точки зрения» (1926). Первым важным теоретическим выводом Гурвича явилось представление о дуализме факторов, вызывающих митоз только при их сочетании. Один из этих факторов (или группа факторов) связан с эндогенными процессами подготовки клетки к делению (фактор возможности или готовности). Другой является экзогенным по отношению к данной клетке (фактор осуществления). Дальнейшие исследования Гурвича были посвящены главным образом изучению второго фактора.

Александр Гаврилович Гурвич. 1874–1954.
Эксперименты и теоретические рассуждения привели Гурвича в 1923 г. к открытию, что большинство экзотермических реакций, как в организме, так и в пробирке сопровождается УФ-излучением. Важнейшим биологическим следствием такого явления оказалась стимуляция клеточных делений, почему эти лучи получили название митогенетических, т. е. вызывающих митозы. В течение последующих лет Гурвичем (1948, 1959) и его сотрудниками было выполнено большое число исследований, посвященных проблеме митогенетического излучения. Стимулирующее влияние излучения было выяснено на самых разнообразных объектах — от бактерий и дрожжевых грибков до зародышей и клеток культуры ткани млекопитающих (А.А. Гурвич, 1968).
В первой четверти XX в. стали накапливаться данные относительно влияния на митоз внешних воздействий — лучистой энергии, различных химических веществ, температуры, концентрации водородных ионов, электрического тока и т. д. Особенно много исследований было выполнено на культуре ткани. В настоящее время установлено, что митотическое деление является следствием длинной цепи причин.
В противоположность цитологии начального периода, которая уделяла основное внимание самому митозу, современная цитология гораздо больше интересуется интерфазой. Пользуясь терминологией Гурвича, можно сказать, что сейчас на первом плане стоит изучение факторов готовности, обеспечивающих возможность вступления клетки в деление. Это стало возможным благодаря новым методам исследования, в первую очередь благодаря радиоавтографии.
А. Говард и С. Пелк (1951) предложили весь митотический цикл разбить на четыре периода: постмитотический, или пресинтетический (G1); синтетический (S), во время которого происходит репликация ДНК; постсинтетический, или премитотический (G2); и, наконец, митоз (М). Накоплен большой фактический материал по продолжительности у самых различных организмов отдельных периодов и всего митотического цикла в целом в норме и при воздействии разнообразных внешних и внутренних факторов — лучистой энергии, вирусов, гормонов и т. д.
Ряд исследований (М. Суонн, 1957, 1958) посвящен энергетике клеточного деления, и хотя многие детали остаются еще невыясненными, стало очевидным, что важная роль принадлежит в этом отношении макроэргическим соединениям, в частности АТФ. Это вещество не только участвует в подготовке клетки к делению, но, по данным Г. Гофман-Берлинга (1959, 1960), ответственно за механические процессы, лежащие в основе расхождения хромосом к полюсам.
В выяснении механизма различных этапов клеточного деления особенно большую роль сыграли работы американского исследователя Д. Мезия (1961), изучавшего различные стороны физиологии митоза, в особенности роль митотического аппарата, осуществляющего самый процесс деления. Созданы различные представления о механизме разделения клеточного тела и о физико-химических изменениях клеток при делении. Изучение хромосом выросло в самостоятельную область исследований, которая оказалась органически связанной с генетикой и дала начало цитогенетике.
Наряду с изучением отдельных митозов значительное число исследований было посвящено выяснению закономерностей митотической активности тканей, в частности изучению зависимости клеточной пролиферации от физиологического состояния организма и влияния различных эндогенных и экзогенных факторов.
Первые исследования такого характера были выполнены на растительных объектах в самом начале XX в. в связи с изучением периодичности биологических процессов (А. Льюис, 1901; В. Келликот, 1904). В 20-х годах появился ряд фундаментальных исследований, посвященных суточному ритму клеточных делений в проростке растений (Р. Фризнер, 1920; М. Столфелд, 1921). В 30-40-х годах была проведена серия исследований (А. Карлетон, 1934; Ч. Влюменфельд, 1938, 1943; 3. Купер, Г. Франклин, 1940; Г. Блюменталь, 1948; и др.), в которых изучалась митотическая активность в очагах клеточного размножения различных лабораторных животных. Значительно меньше таких работ выполнено на очагах клеточного размножения человека (3. Купер, А. Шифф, 1938; А. Бродерс, В. Дублин, 1939; и др.).
В СССР первое исследование по влиянию на митотический режим физиологических факторов было опубликовано в 1947 г. Г.К. Хрущовым. Начиная с 50-х годов интерес к проблеме митотического режима организма значительно возрос (С.Я. Залкинд, И.А. Уткин, 1951; С.Я. Залкинд, 19,54, 1966; В.Н. Доброхотов, 1963; И.А. Алов, 1964; и др.). Наиболее полно был изучен суточный ритм митотической активности у млекопитающих.
Первые попытки проанализировать механизмы, регулирующие митотическую активность, были предприняты в 1948 г. английским исследователем В. Буллоу. Советские цитологи (Л.Я. Бляхер, 1954; И.А. Уткин, 1959; Г.С. Стрелин, В.В. Козлов, 1959) уделили большое внимание нейрогуморальной регуляции митотической активности, установив ?рефлекторный характер регуляции клеточных делений. Оказалось, что воздействие на нервную систему влияет опосредованно — через сдвиг гормонального равновесия. Выяснилось также, что при этом резко усиливается секреция адреналина, тормозящего митотическую активность. Удаление надпочечников приводит к выключению эффекта торможения митозов (А.К. Рябуха, 1955, 1958). Ряд исследований посвящен изучению сложных взаимоотношений между митотической и физиологической активностью организма (С.Я. Залкинд, 1952; И.А. Алов, 1964).
Повышение интереса к проблеме митотических циклов и широкое применение радиоавтографии привело к тому, что в настоящее время подавляющее большинство работ посвящено изучению закономерностей митотического цикла, анализу закономерностей перехода из одного периода в другой, влиянию на митоз разнообразных эндогенных и экзогенных факторов. Это, несомненно, одно из наиболее перспективных направлений в изучении проблемы клеточной пролиферации (О.И. Епифанова, 1973).
Цитология наследственности.
В первой половине XX в. в связи с расцветом генетики интенсивно разрабатывались цитологические проблемы, касающиеся наследственности. Так возникла новая область цитологии — кариология.
Пионером кариологических исследований был русский ботаник С.Г. Навашин. Навашин по справедливости может быть назван создателем цитогенетики, не случайно первый период в развитии этой науки часто называют «русским» или «навашинским». Уже в классических работах по эмбриологии растений, в особенности по цитологии оплодотворения (1898), он сосредоточил свое внимание на морфологии хромосом в клетках некоторых лилейных, в частности, конского гиацинта (Galtonia candicans). В 1916 г. Навашин опубликовал работу, в которой привел тщательное описание хромосомного набора этого растения. Ему удалось найти на хромосоме (в центре или на ее полюсе) особый неокрашенный участок (названный им «хроматическим перерывом»), именуемый сейчас центромерой или кинетохором, в области которого хромосома прикрепляется к веретену. Центромерам принадлежит чрезвычайно важная роль в процессе расщепления хромосом и их расхождения к полюсам делящейся клетки. Навашин впервые показал, что строение хромосом вовсе не является неизменным, но подвержено изменениям в филогенезе и при некоторых особых условиях существования (например, в клетках семян при их длительном хранении). На ряде растительных объектов (Crepis, Vicia, Muscari и др.) ученики Навашина показали, что кариолотический анализ может быть использован для филогенетических выводов. Несколько позже начались кариологические исследования на клетках животных и человека. В этих работах также участвовал Навашин. Уже после его смерти, в 1936 г., была опубликована работа, посвященная уменьшению (диминуции) хроматина при развитии яйца лошадиной аскариды, подтвердившая выводы Т. Бовери (1910).
Обстоятельные кариологические работы были выполнены в 20-30-х годах советским цитологом П.И. Живаго. Он и его сотрудники исследовали кариотип домашних птиц (куры, индейки; 1924, 1928), мелкого рогатого скота (1930) и человека (1932). Живаго не только выяснил ряд кариотипов, но и начал разработку вопроса о постоянстве числа хромосом в пределах одного организма. На основании литературных данных (по двукрылым) и исследования ряда объектов (эму, нанду, человек) Живаго (1934) пришел к заключению, что в отдельных клетках и целых тканях (особенно у эмбрионов) наблюдаются значительные колебания в числе хромосом. Он придавал этим различиям большое значение, так как они ведут к изменению генома, а следовательно, и наследственных свойств организма. Он высказывал также предположение, что наличие клеток с различным числом хромосом может иметь приспособительное значение, так как увеличивает возможные- варианты кариотипов для последующего отбора. Эта точка зрения, высказанная свыше 30 лет тому назад, разделяется в настоящее время многими исследователями.
Большую роль в развитии рассматриваемого направления сыграла книга К. Белара «Цитологические основы наследственности» (1928, русский перевод 1934). Разделу, посвященному связи хромосом с наследственностью, предшествуют собственно цитологические главы, содержащие данные о строении ядра и цитоплазмы, о клеточном делении, оплодотворении и созревании половых клеток, о партеногенезе. Очень детально и в сравнительном аспекте рассматривается строение хромосом не только у высших позвоночных, но и у беспозвоночных, простейших и растений. Содержатся ценные данные, касающиеся индивидуальности и изменчивости хромосом, обмена фрагментами при кроссинговере, диминуции хроматина, патологии митоза. Книга Белара в течение долгого времени оставалась лучшей монографией по цитологии наследственности.
Постепенно, в связи с интенсивным развитием генетики, цитология наследственности превратилась в цитогенетику, история которой кратко изложена вместе с историей генетики (см. главы 13 и 24).
Новейшие направления в цитологии.
Во второй половине XX в. возникло несколько совершенно новых, весьма перспективных направлений исследований.
В первую очередь следует назвать цитоэкологию, изучающую роль клеточного уровня организации в приспособлении организма к условиям среды. В СССР это направление, тесно связанное с биохимией клетки и особенно с изучением свойств клеточных белков, получило широкое развитие в работах В.Я. Александрова и Б.П. Ушакова.
За последние 10–20 лет большое внимание привлекает изучение общей физиологии клетки и, в частности, закономерностей синтеза и расходования веществ, как участвующих в основных жизненных процессах, так и являющихся ее специфическими продуктами (секреты). К этому же кругу вопросов относится изучение восстановительных процессов в клетке, т. е. физиологической регенерации, обеспечивающей восстановление разрушенных или утраченных клеточных структур и веществ и совершающейся на молекулярном уровне.
Большое значение в цитологии приобрели проблемы детерминации, дифференциации и дедифференциации клеток. Они играют важную роль в эмбриональных клетках и различных категориях клеток, культивируемых вне организма (А. Де-Рейк, Дж. Найт, 1967; С.Я. Залкинд, Г.Б. Юровская, 1970).
Своеобразный раздел цитологии составила цитопатология — область, пограничная с общей патологией и сделавшая значительные успехи в последние десятилетия XX в. Термин «цитопатология» используется для обозначения отрасли биологии, в которой изучение общепатологических процессов ведется на клеточном уровне, и как система знаний о патологических изменениях отдельной клетки. Что касается первого направления, то после классических работ Р. Вирхова попытки свести сущность патологического процесса к изменению микроскопических и субмикроскопических структур предпринимались неоднократно. Много примеров подобного использования цитологического анализа для понимания патологических процессов в организме содержится в работах Р. Камерона (1956, 1959).
Второе направление может рассматриваться как чисто цитологическое. Оно ставит своей целью изучение патологии самой клетки и ее органоидов, т. е. морфологических, биохимических и физиологических отклонений от нормы, наблюдаемых при происходящих в клетке различных патологических процессах, независимо от их влияния на состояние ткани, органа или всего организма. Развитие этого направления связано прежде всего с накоплением данных об изменении клеток, происходящем вследствие их естественного старения, а также различных резких цитопатологических изменений, наблюдаемых при воздействии тех или иных неблагоприятных факторов (физических, химических, биологических) внешней среды. Особенно значительное развитие получило изучение патологических изменений под влиянием неблагоприятных воздействий на клетку в эксперименте и исследование механизма действия таких факторов. Эти исследования получили широкое развитие в первую очередь в радиобиологии, где всестороннее изучение реакции клетки на воздействие лучистой энергии возможно не только на клеточном или субклеточном, но и на молекулярном уровне.
Значительное развитие получила вирусная цитопатология, т. е. изучение реакции клетки (чаще всего клеток однослойных культур) на воздействие вируса. При этом удается не только проследить все этапы воздействия вируса, но и, пользуясь разнообразными методами современного исследования, выяснить реакцию самой клетки на вирусную инфекцию. Эти исследования имеют не только большое теоретическое значение (для решения проблемы взаимоотношения вируса и клетки), но важны и в практическом отношении, так как по характеру специфического действия вируса на клетки может быть с достаточной степенью достоверности установлена природа вируса, воздействующего в каждом конкретном случае. Диагностика вирусов по вызываемому ими эффекту является одним из наиболее распространенных методов прикладной вирусологии.
Несомненно, перспективным является изучение разных проявлений патологии митоза — повреждения хромосом, митотического аппарата, нарушения разделения клеточного тела (цитотомии). Ведущиеся в этом направлении исследования призваны помочь вскрыть не только нормальные механизмы этого важнейшего общебиологического процесса, но и некоторые стороны канцерогенеза и особенности раковых клеток. Возможно, здесь будет открыт путь к возвращению таких клеток в нормальное состояние.
В настоящее время имеются, например, данные, что для обеспечения правильного осуществления митоза особое значение имеет сульфгидрильно-дисульфидное равновесие в клетке; его нарушение может самым существенным образом отразиться на формировании митотического аппарата, обеспечивающего нормальное протекание этого процесса (И.А. Алов, 1966).
Особенности современной цитологии.
Если коротко охарактеризовать особенности современной цитологии, то следует прежде всего отметить, что эта область знания проделала в XX в. значительную эволюцию. В начале века в цитологию стали проникать экспериментальные методы исследования. Направление науки в этот период можно назвать аналитическим. Основным содержанием работ был анализ физико-химических и физиологических свойств клетки.
В 40-50-х годах XX в. положение изменилось — цитология превратилась в клеточную биологию. Ее содержанием стало изучение основных общебиологических вопросов на микроскопическом, субмикроскопическом (электронно-микроскопическом), макромолекулярном и молекулярном уровнях.
В центре внимания современной цитологии оказались не только структуры клетки, но в еще большей степени те вещества, из которых они построены. В равной мере преимущественный интерес вызывают наиболее общие процессы, протекающие в клетке и создающие основу ее жизнедеятельности (передача информации от ДНК к РНК, синтез белка, генерация энергии, активный транспорт веществ и т. д.). Проблемы классической цитологии — митоз, мейоз, секреция, проницаемость и т. д. — изучаются в настоящее время в свете данных об элементарных процессах, происходящих в клетке, и задачей соответствующих исследований чаще всего оказывается не феноменологическое описание, а выяснение механизмов основных проявлений жизнедеятельности. Ярким подтверждением актуальности этого нового направления цитологии является не только то, что «Биологией клетки» назван один из наиболее распространенных учебников цитологии Э. Де-Робертиса, Ф. Caeca и В. Новинского[89], но и тот факт, что на протяжении последних 20 лет международные конгрессы цитологов именуются конгрессами по биологии клетки.
Развитие современной цитологии находит свое выражение и в некоторых научно-организационных формах, в частности в создании обществ, изучающих отдельные разделы цитологии. Помимо международного объединения исследователей, работающих в области биологии клетки, можно назвать объединения цитохимиков, электронных микроскопистов, исследователей, входящих в группу по изучению закономерностей клеточной пролиферации и т. д. Выходит большое число журналов, специально посвященных проблемам цитологии. В СССР организованы специальные институты цитологии (в Ленинграде и Новосибирске); цитологическая тематика разрабатывается в ряде институтов Академии наук и Академии медицинских наук СССР (Институты биологии развития, морфологии человека, медицинской генетики и др.) и институтов союзных республик. Цитологические работы публикуются в ежемесячном журнале «Цитология». Работу в области цитологии планирует и координирует Научный совет по проблемам цитологии при АН СССР.
Наряду с обособлением и растущей дифференциацией проблематики цитология вступает во все более тесные связи со смежными биологическими дисциплинами. Достижения современной цитологии широко используются вирусологией, микробиологией, иммунологией, онкологией и др. Многие общебиологические и медицинские вопросы разрешаются сейчас на клеточном уровне.
Некоторые перспективы развития цитологии.
Оценивая ближайшие перспективы развития цитологии, следует сказать прежде всего о будущем аналитического направления исследований. Оно, очевидно, приобретает еще более углубленный характер. Потребуется более детальное, комплексное (электронно-микроскопическое, био- и гистохимическое) исследование всех структурных компонентов клетки. Будет изучаться биологическая роль макромолекул в клетке, так как именно макромолекулы являются основными элементами, определяющими как структуры клетки, так и механизмы процессов ее жизнедеятельности.
Можно предполагать, что при этом еще большее развитие получат наиболее тонкие и точные методы инструментального исследования и дифференциальный анализ компонентов клетки будет доведен до высокой степени совершенства. На этом пути цитология в известной мере сольется с молекулярной биологией, которая уже и сейчас глубоко проникла в цитологию. Границы последней в этом случае будут определяться объектом — клеткой и теми специальными задачами, которые могут быть поставлены при таком углубленном анализе.
Однако наряду с аналитическим направлением в цитологии будущего, может быть, еще большую роль приобретет направление, которое можно назвать синтетическим, или интегративным. Уже сейчас в цитологии важное место занимают такие проблемы, как детерминация, дифференциация, синхронизация клеточного размножения, гибридизация и генетика соматических клеток, вопросы клеточных взаимодействий и контактов. Нет сомнения, что все эти вопросы в будущем получат еще более интенсивное развитие.
Сейчас в цитологии все большее значение приобретает изучение механизмов контроля, регуляции и интеграции различных сторон деятельности не только самой клетки (например, регуляция клеточного размножения), но и отдельных ее органоидов. По-прежнему будет развиваться изучение роли клетки в составе систем тканей и органов. Результатом этих исследований явятся более полные синтетические данные о различных сторонах жизни клеток и закономерностях их взаимодействия.
Важное место займет изучение проблемы специфичности клетки в двух аспектах — как ее ответ на внешние воздействия и как генетически обусловленная детерминация. Уже имеются данные, свидетельствующие о том, что реакция клетки на воздействие внешних факторов во многом зависит от способности клетки к адаптации, ее консерватизма, способности к перестройке и т. д. Изучение конкретных механизмов этой специфической реакции составит, очевидно, одно из основных направлении цитологии ближайшего будущего. Имеются данные, касающиеся «иммунологической памяти» по крайней мере некоторых клеточных категории, что позволяет говорить о способности клеток к «узнаванию» иммунологически подобных элементов. Вскрыть механизм этого своеобразного свойства — также одна из ближайших задач цитологии.
Важным и перспективным направлением цитологии является изучение реактивности клеток. Этот вопрос имеет не только теоретическое, но и существенное практическое значение в связи с воздействием на клетку различных повреждающих факторов, установлением пределов и условий ее репарации и т. д. Полученные в этой области данные позволяют во многих случаях говорить о неспецифической устойчивости клетки к различным повреждающим воздействиям. В основе этой устойчивости лежат, очевидно, определенные изменения биосинтетических процессов (А.А. Ябров, 1967).
Таким образом, наряду с аналитическими в цитологии будут, очевидно, развиваться исследования, направленные на изучение механизмов саморегуляции клетки как автономной биологическои системы. Эти вопросы имеют прямое отношение к одной из наиболее трудных и вместе с тем наиболее важных проблем цитологии будущего — проблеме целостности клетки.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК