Глава 22 Проблема возникновения жизни на Земле
Проблема возникновения жизни на нашей планете, а может быть и на других небесных телах, встала перед человеком с той поры, как он начал познавать себя и окружающий мир. За многовековую историю человечества была выдвинута не одна гипотеза о путях возникновения жизни и о месте человека в системе живых существ. Однако только около ста лет назад, после создания Ч. Дарвином теории происхождения видов и особенно появления теории диалектического материализма, проблема происхождения жизни вошла в рамки рациональной науки.
Первые гипотезы.
Фундаментальные положения диалектического материализма о жизни как особой форме движения материи открыли методологические подходы к разрешению проблемы возникновения жизни.
В конце прошлого и начале нашего века в представлениях о происхождении жизни господствовало механистическое метафизическое направление, сторонники которого считали, что происхождение жизни может быть понято только в результате обнаружения в природе самозарождения. Наряду с такой точкой зрения широкое распространение имела теория панспермии С. Аррениуса (1895; лауреат Нобелевской премии, 1903) о заносе на Землю живых существ из космического пространства. При этом вопрос о первичном возникновении жизни на космических телах не затрагивался.
Особенно широкое хождение имели предположения о заносе зародышей живых организмов на Землю с метеоритами или с космической пылью. Однако все попытки обнаружить в метеоритах какие-либо признаки живого не увенчались успехом. Было убедительно показано, что обнаруживаемые в метеоритах микроорганизмы попадали туда при падении метеорита на Землю. В недавнее время американский исследователь Б. Наги (1962) и другие сообщили об обнаружении ими в углистых хондритах «оргей» остатков организмов, якобы обитавших в прошлом на метеоритном материале. Но вскоре было показано, что найденные структурированные образования фактически являются минеральными гранулами и лишь по виду напоминают биологические структуры.
Некоторые биологи в нашей стране длительное время также находились под влиянием взглядов С. Аррениуса. Так, В.И. Вернадский (1939), говоря о несостоятельности теории самозарождения, пришел к выводу, что жизнь должна была когда-то иметь другое начало, отличное от того, которое мы наблюдаем сейчас при зарождении живого организма из такого же живого. По его мнению, историю жизни следует искать не в геологической, а в космической истории Земли. Позже Вернадский уже не говорил о непреодолимой пропасти, разделяющей живую и неживую материю, а лишь отмечал их различный изотопный состав. В 1944 г. он писал: «В наше время этот вопрос едва ли мог так просто трактоваться, как это было возможно в прошлом столетии, когда вопрос об абиогенезе, казалось, был решен окончательно в отрицательном смысле после работ Л. Пастера…»[174].
A.В. Немилов утверждал, что жизнь существовала во Вселенной и тогда, когда еще не было земного шара; он нацело исключал возможность возникновения жизни на Земле из неорганической материи, полагая, что она занесена на Землю с другого космического тела. Теорию панспермии разделяли также С.П. Костычев, П.П. Лазарев и другие.
B.И. Ленин в ряде философских произведений с позиций диалектического материализма обобщил важнейшие открытия естествознания того времени. Особо важное значение для решения проблемы происхождения жизни имело положение Ленина о важнейшей роли экспериментальных исследований в материалистической разработке нерешенных проблем естествознания.
Значение книги А.И. Опарина (1924).
Начало систематической разработке проблемы возникновения жизни на Земле было положено в 1924 г. в связи с выходом в свет книги «Происхождение жизни» А.И. Опарина[175]. Здесь он впервые в общих чертах попытался сформулировать естественнонаучную теорию возникновения жизни на Земле как результат длительного эволюционного развития материи. Диалектический принцип развития, положенный в основу философского анализа проблемы, позволил Опарину связать рассматриваемый вопрос с проблемой сущности жизни. Обобщив накопленный естествознанием фактический материал, он проследил в естественно-историческом аспекте образование и последующую эволюцию простейших структур, энергетических процессов и биохимических функций, которые могли иметь место на Земле в период возникновения и становления, жизни. Как отмечал Дж. Бернал, эта теория легла в основу почти всех современных представлений о возникновении жизни. В 1929 г. была опубликована статья Дж. Холдейна, в которой он также развивал эволюционные взгляды на проблему возникновения жизни. Но как позже отмечал сам автор, в его статье вряд ли можно было найти что-либо новое, чего бы не было в книге Опарина. В последующие годы теория Опарина в основных своих положениях получила полное подтверждение. Как отметил Н. Горовиц (1957), огромное достоинство теории состоит в том, что большая ее часть может быть проверена или логически связана с поддающимися проверке предположениями. Именно это обстоятельство побуждало многих исследователей различных областей современного естествознания к экспериментальному воспроизведению описанных процессов, происходивших на разных этапах развития материи.
Первичное образование на Земле простейших органических веществ.
Правильный научный путь познания последовательных ступеней развития материи, приведшего к возникновению жизни на нашей планете, состоял в изучении процессов первичного образования простейших органических веществ, без которых жизнь невозможна. Было выяснено, что первым, чрезвычайно важным шагом на пути к возникновению жизни был переход неорганических соединений углерода в органические.
Еще в начале XX в. большинство ученых считало, что в природных условиях органические вещества образуются только в результате деятельности живых существ. К.А. Тимирязев, например, указывал, что единственной лабораторией, где заготовляется органическое вещество на оба царства природы, является зеленый лист. Другим биогенным источником органического вещества является хемосинтез, открытый С.Н. Виноградским (1887).
Большинство ученых считало первичные организмы автотрофами, т. е. способными питаться неорганическими веществами; органические же вещества на Земле имеют якобы вторичное происхождение. Эти представления составили главное препятствие, ставшее на пути любых попыток понять пути эволюции материи, приведшей к появлению жизни.
В 1922 г. Опарин высказал предположение, что органические вещества на нашей планете должны были образовываться абиогенным путем задолго до появления на ней жизни. Вместе с тем он полагал, что первичные живые существа могли быть и гетеротрофами, т. е. были способны питаться уже готовыми органическими веществами. Этот вывод вытекал из данных о составе звездных атмосфер, а также метеоритов, в которых удалось обнаружить присутствие углеводородов. В последнее время в составе метеоритов особое внимание привлекли углистые хондриты, содержащие значительные количества углерода, гидратированные силикаты и алюмосиликаты. Наряду с низкомолекулярными углеводородами в углистых хондритах, как показал Г.П. Вдовыкин (1960, 1963), содержатся также высокомолекулярные полимеризованные соединения, жирные кислоты и органические вещества ароматического ряда.
Следует отметить, что предположения о возможности возникновения жизни путем эволюционного развития материи высказывались еще К.А. Тимирязевым, Э. Пфлюгером, Дж. Тиндалем и А. Шефером. Однако из-за господствовавших взглядов о биогенном происхождении органического вещества эти представления не получили в то время развития. Заслуга Опарина состоит в том, что он впервые указал на ограниченность этих взглядов, основанных лишь на учете тех условий, которые существуют на Земле в настоящее время. Данные астрономии, космогонии, геофизики и астрофизики показывают, что и сейчас повсеместно происходит образование органических веществ совершенно независимо от жизни. Имеются факты, свидетельствующие о том, что при формировании земной коры должны были возникать углеводороды (как в газообразном, так и в жидком состоянии), и уже в газопылевой материи, из которой образовалась наша планета, должны были присутствовать органические соединения различной сложности. Таким образом, органический синтез, вероятно, осуществлялся и в период, предшествовавший образованию солнечной системы, и во время ее образования. Ученые пришли к заключению, что такой синтез проходил в атмосфере углеродных звезд, в солнечной туманности, протопланетах и планетозималиях (кремневые и железные тела, подобные современным метеоритам). Точку зрения о существовании органических веществ до возникновения нашей планеты разделял также Дж. Бернал.
В последнее время Дж. Бернал (1967) выдвинул гипотезу космического происхождения органических веществ. Она опирается на данные исследования органических веществ углистых хондритов и метеоритной пыли красных океанических глин. Однако американские исследователи С. Миллер и Г. Юри (1953) считают, что органические вещества, синтезированные еще до образования Земли, не могли играть решающей роли в процессе возникновения на ней жизни. По их мнению, доля органических соединений, занесенных на Землю с кометным материалом, очень мала. По-видимому, основная масса органических веществ возникла на Земле при образовании земной коры. Абиогенное образование углеводородов и циана, явившееся первой ступенью в развитии органической материи, в настоящее время не вызывает никаких сомнений, поскольку эти углеродные соединения удается обнаружить на всех небесных телах, доступных изучению.
Восстановительный характер первичной земной атмосферы.
Условия, некогда существовавшие на еще безжизненной Земле, во многом отличались от современных. Это касается прежде всего состава первичной земной атмосферы. В.И. Вернадский (1926) впервые указал, что подавляющая масса газов, образующих земную атмосферу, имеет биогенное происхождение. На первых порах в атмосфере Земли отсутствовал свободный кислород, что обеспечивало беспрепятственный доступ к земной поверхности ультрафиолетового излучения. В сравнении с современными условиями это создавало гораздо большие возможности для разнообразных фотохимических процессов. Нынешние атмосфера, верхняя часть почвы и вся гидросфера богаты свободным кислородом и густо населены микроорганизмами. Последние, потребляя органические вещества внешней среды, исключают возможность их длительной эволюции. Поэтому при решении проблемы возникновения жизни задача исследователей состоит в том, чтобы попытаться воссоздать в лаборатории те условия, которые должны были существовать на первичной Земле. Экспериментальные исследования в этом направлении уже дали ценный материал по абиогенному синтезу ряда биологически важных соединений.
Для формирования теории возникновения жизни на Земле особое значение имело предположение Опарина о том, что первичная земная атмосфера находилась в восстановленном состоянии, и что на определенном этапе ее развития она должна была содержать наряду с газообразным водородом и парами воды соединения углерода (в виде метана и циана) и азота (в виде аммиака). На эту мысль его натолкнула гипотеза Д.И. Менделеева о неорганическом происхождении нефти, а также обнаружение метана в атмосфере больших планет. Основываясь на новейших астрономических данных и изучении метеоритов, Г. Юри (1952) также пришел к выводу, что первичная атмосфера Земли вследствие значительного содержания в ней водорода носила восстановительный характер. Опарин и Юри полагают, что с течением времени состав атмосферы постепенно изменился: в ней возросло содержание кислорода, и она приобрела окислительный характер.
По мнению голландского геолога М. Руттена (1962), кислород в атмосфере Земли начал появляться около двух миллиардов лет тому назад, но только за миллиард лет до настоящего времени создалась современная окислительная атмосфера. Между этими рубежами лежал, по его мнению, переходный период, когда атмосфера Земли постепенно обогащалась свободным кислородом в результате возникновения начальных форм жизни. К. Саган (1961) считает, что признаки окислительной атмосферы появились значительно раньше.
Современные астрономические данные и изучение метеоритов показывают, что на последних этапах эволюции солнечной системы большинство ее тел характеризовалось восстановительной бескислородной атмосферой, наличием свободной воды, щелочным или нейтральным pH среды, умеренной температурой и сравнительно высокой концентрацией органических веществ. По мнению Дж. Оро (1965), образование органических соединений во Вселенной происходило в результате воздействия тепловой энергии, энергии ионизирующего и ультрафиолетового излучений, а также электрических разрядов. Первичным источником этих форм энергии служат термоядерные процессы, протекающие в недрах звезд. В абиотическом образовании простых органических соединений, имеющих биохимическое значение, принимали, по-видимому, участие следующие источники и механизмы: во-первых, радикалы (в случае синтеза предшественников альфа-аминокислот); во-вторых, активно протекающая в щелочной среде реакция альдольной конденсации простых альдегидов; в-третьих, активно протекающая в щелочной среде конденсация цианистого водорода и других нитрилов и последующий синтез пуринов, пиримидинов и других гетероциклических соединений. При этом, по-видимому, также имели место каталитические процессы и использовались другие внутренние источники энергии.
Абиогенный синтез важнейших органических соединений.
Представления о химической эволюции вещества на пути к возникновению жизни сложились в результате целого ряда экспериментальных работ, в которых были осуществлены синтезы важнейших органических соединений в системах, моделирующих химический состав первичной земной атмосферы. Эти работы служат основным доказательством правомерности теории возникновения жизни, выдвинутой советскими учеными.
Начало серии работ по абиогенному синтезу было положено С. Миллером (1953), синтезировавшим ряд аминокислот при пропускании электрического разряда через смесь газов, предположительно составлявших первичную земную атмосферу (водород, пары воды, метан, аммиак). T.Е. Павловская и А.Г. Пасынский (1957) заменили водород в реакционной смеси на окись углерода. В этом случае при воздействии электрического разряда и ультрафиолетового излучения также происходило образование аминокислот. Очевидно, аминокислоты могли абиогенно образовываться и на той стадии развития Земли, когда значительная часть водорода была утеряна, и атмосфера, сохраняя восстановительный характер, могла содержать большие количества окиси углерода.
П. Эйбелсон (1956), используя газовую смесь, содержавшую наряду с метаном, аммиаком, водородом, парами воды также окись углерода, углекислый газ и азот, получил аналогичную смесь аминокислот. Он показал при этом, что аммиак может быть заменен на азот, а метан на углекислый газ. К. Хейпс (1957) и другие вводили в смесь исходных газов сероводород, но им удалось синтезировать не аминокислоты, содержащие серу, а лишь тиоацетат и тиомочевину.
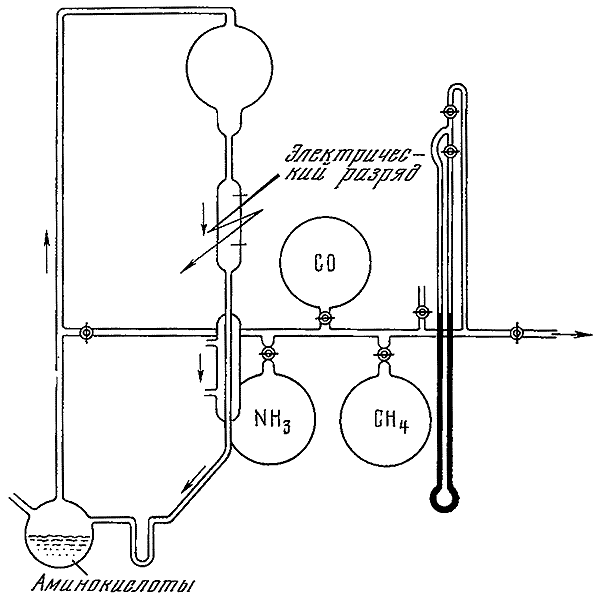
Прибор С. Миллера (1955) для синтеза аминокислот в электрическом разряде.
К. Дозе и Б. Раевский (1957) обнаружили образование кислых и нейтральных аминокислот при действии рентгеновых лучей на смесь первичных газов. Т. Гассельстромом и другими исследователями (1957) были получены глицин и аспарагиновая кислота при облучении бета-лучами растворов ацетата аммония. П. Пашке (1957) наблюдал образование глицина и аланина при облучении твердого карбоната аммония гамма-лучами. А.Г. Пасынский и Т.Е. Павловская (1959) показали возможность образования аминокислот при действии ультрафиолетового излучения на раствор смеси формальдегида и солей аммония. К. Гроссенбахер (1963), пропуская длительное время искровой разряд через смесь аммиака, метана и водорода, обнаружил наряду с аминокислотами также пептиды, которые к концу опыта выделились из раствора в виде мелких сферических образований или капель. И.Е. Эльпинер (1960–1970) осуществил синтез ряда аминокислот и пептидов при действии ультразвуковых колебаний на воду, насыщенную водородом, азотом и окисью углерода, а также синтез имидазола. Дж. Оро (1963) произвел абиогенный синтез аденина, гуанина, пиримидинов, рибозы и дезоксирибозы. С. Поннамперума (1963) для синтеза нуклеотидов в водной среде применил этилметафосфат, а в качестве источника энергии — ультрафиолетовое излучение с длиной волны 2400–2900 А, которое, по-видимому, получала поверхность первичной Земли. При этом ему удалось превратить аденин в аденозин, аденозин в аденозиномонофосфат, а последний — в аденозинтрифосфат. Таким образом, он показал возможность абиогенного синтеза АТФ — основного источника энергии живых организмов. А.А. Красновский и А.Г. Умрихина (1964) показали, что в присутствии кислорода облегчается синтез порфирина из пиррола и формальдегида. А. Шутка (1959) осуществил синтез порфирина и его производных абиогенным путем в присутствии кислорода. Это дало ему основание сделать вывод, что эти соединения возникли сравнительно поздно, когда в земной атмосфере появился кислород.
Существенным этапом в разработке рассматриваемой проблемы явился первый Международный симпозиум по происхождению жизни, состоявшийся в Москве в 1957 г. На этом симпозиуме А.Н. Белозерский высказал мысль о том, что в абиогенном синтезе определенную роль могли играть неорганические полифосфаты. В 1958 г. Г. Шраммом была осуществлена поликонденсация различных биологически важных соединений в безводной среде при температурах +55–60 °C с применением эстерифицированных полифосфатов. Им были получены некоторые пептиды, полисахариды, нуклеотиды и нуклеозиды, а также полиаденин и полиурацил. Это дало ему основание высказать предположение, что на поверхности Земли белковоподобные и нуклеиноподобные полимеры могли синтезироваться при помощи полифосфатов, которые легко вовлекались в простейшие механизмы сопряжения химических реакций, связанные с возникновением жизни. В 1963 г. на симпозиуме в США, посвященном проблеме происхождения предбиологических систем, С. Фокс с сотрудниками сообщили о работах по синтезу аминокислот из метана, аммиака и паров воды в лабораторных условиях, близких к вулканическим. Так как этот вид энергии на первичной Земле носил, по-видимому, локальный характер, термический синтез мог иметь определенное значение для абиогенного образования аминокислот. При температурах 150–180 °C ими была осуществлена также термическая поликонденсация различных аминокислот, в результате которой образовались полипептиды с молекулярным весом 3000–9000. По целому ряду показателей протеиноиды Фокса не отличаются от таких белков, как казеин. Они выделяются при помощи 1 %-ного водного раствора поваренной соли в виде обнаруживаемых под микроскопом шариков-микросфер.
Дальнейшим важным шагом эволюции органических веществ на пути к возникновению жизни была полимеризация мономерных компонентов и образованием первичных полипептидов и полинуклеотидов.
Возникновение предбиологических систем.
Приведенные данные по абиогенному синтезу важных биохимических соединений подверглись обсуждению на упоминавшемся симпозиуме в США (1963). Было отмечено, что теория возникновения жизни, разрабатываемая советскими учеными, стимулировала экспериментальные и теоретические исследования в области абиогенного синтеза органических веществ и привела к крупным достижениям в этой области. Участники симпозиума выразили уверенность, что хотя начальные этапы синтеза многих биологически важных молекул еще не воспроизведены, есть все основания рассчитывать, что их удастся воссоздать в ближайшем будущем.
В настоящее время считается установленным, что Земля существует около 4,5 млрд. лет, а жизнь появилась на ней примерно 3 млрд. лет тому назад. Следовательно, на протяжении значительной части времени существования Земли она оставалась безжизненной. В атмосфере и гидросфере Земли находилось достаточное количество углеводородов, которые и послужили исходным материалом для последующей химической эволюции. Основная их масса возникла при формировании земной коры, незначительная часть была занесена с кометным и метеоритным материалом. Согласно подсчетам Г. Юри (1952) и К. Сагана (1961), за период в миллиард лет концентрация органических веществ, синтезированных в атмосфере и оседавших в водах Мирового океана, должна была достигнуть 1 %. Таким образом, на определенном этапе существования Земли эти воды превратились в Своеобразный «первичный бульон», содержавший наряду с неорганическими солями также и разнообразные органические вещества.
Особое значение имеет решение вопроса о том, как в условиях первичной Земли могли образоваться белковоподобные и нуклеиноподобные вещества, а также ферменты, являющиеся основными участниками, проводниками и регуляторами всех жизненных процессов. В связи с тем, что путем абиогенного синтеза получены не все аминокислоты, входящие в состав современных белков, было высказано предположение, что некоторые аминокислоты возникли на более поздних ступенях развития органического вещества. Их отсутствие могло ограничивать каталитические функции первичных белков.
В этой связи большой интерес представляют исследования японского биохимика Ш. Акабори (1955), высказавшего предположение, что первичные белки не обязательно должны были образовываться из готовых аминокислот. Лабораторными опытами Акабори показал, что образование «предбелков» могло происходить и из таких органических соединений, как формальдегид, аммиак и цианистый водород. Продукты, образующиеся при их взаимодействии, полимеризуются затем на поверхности глины, после чего в результате гидролиза могут образоваться полипептиды, в частности, полиглицин.
Характерной чертой живых существ является синтез асимметрических органических соединений. Так, Акабори (1962) осуществил асимметрическое гидрирование карбонильных соединений на никелевом катализаторе. А.Н. Терентьев и Е.М. Клабуновский (1957), рассматривая вопрос о роли асимметрии в возникновении жизни, пришли к мысли, что асимметрия органического мира является следствием воздействия циркулярно поляризованного света и асимметрической решетки кристаллов. По их мнению, присутствие асимметрических органических катализаторов направляло реакции в сторону образования оптически чистых продуктов. Согласно Г. Уолду (1957), геохимические синтезы органических веществ могли производить лишь рацемические смеси. Отбор отдельных оптических изомеров происходил при образовании структур более высокого порядка — полипептидов, белков и нуклеиновых кислот.
Эволюция предбиологических систем.
Следующей загадкой был вопрос, каким образом в «первичном бульоне»- возникли первые живые организмы, обладавшие обменом веществ и способностью к самовоспроизведению. Иначе говоря, как из хаоса бесконечного числа допустимых термодинамикой перекрещивающихся химических реакций возник упорядоченный обмен веществ, обеспечивающий сохранение и развитие живой системы?
По этому вопросу существуют разные точки зрения. Согласно одной из них, разделяемой Н. Горовицем и У. Стенли[176], появление первых организмов шло путем молекулярной эволюции первоначально возникших молекул со случайным расположением мономеров в направлении образования молекул с их упорядоченной последовательностью. По этим представлениям, «началом жизни» явилась молекула нуклеиновой кислоты или нуклеопротеида (вирус). В пользу этих представлений свидетельствовали, в частности, данные А. Корнберга (1961)[177] о том, что ДНК при определенных условиях может удваиваться и вне клетки. Как показали, однако, дальнейшие исследования, такое удвоение возможно лишь в том случае, если в инкубационной среде присутствуют различные сложные компоненты клетки, существование которых в первичном океане трудно себе представить. Обсуждая эти представления, Опарин подчеркивал, что характерным для жизни является то, что она не просто рассеяна в пространстве, а представлена отграниченными от внешнего мира индивидуальными системами-организмами. Последние могли возникнуть только на основе длительной эволюции, постепенного совершенствования каких-то гораздо более простых исходных систем, выделившихся из первичного однородного «бульона». Поэтому едва ли можно полагать, что в процессе эволюции сперва возникли белки, нуклеиновые кислоты и другие вещества протоплазмы, строение которых было чрезвычайно хорошо приспособлено к выполнению определенных биологических функций, а затем на основе их объединения возникла сама живая протоплазма. В условиях первичной Земли появление свойственной живым существам организации могло произойти только на базе эволюции хотя и более примитивных, но целостных многомолекулярных систем. В момент своего образования эти системы еще не обладали специфическими для жизни особенностями; их превращение в специфические системы более высокого порядка, характерные для живых организмов, есть результат длительной эволюции на основе естественного отбора. Возникшая в дальнейшем на основе формирования генетического кода способность к передаче наследственной информации от предков к потомкам стала одним из основных свойств организмов. Эта точка зрения находит в наши дни все больше сторонников.
Коацерватные капли как модель предбиологической системы.
В 1924 г. в первом наброске своей теории, Опарин высказал предположение, что образующиеся при смешении растворов различных белков и других высокомолекулярных веществ коллоидные гели могли явиться формой организации многомолекулярных систем и стать объектом эволюции, приведшей к возникновению жизни. Явление отслаивания коллоидных гелей было названо голландским исследователем Г. Бунгенберг-де-Ионгом коацервацией. Образующиеся в результате этого процесса коацерватные капли были подвергнуты им и его школой коллоидно-химическим исследованиям.
За последние годы главным образом в Институте биохимии им. А.Н. Баха Академии наук СССР и в Московском государственном университете детально изучен химический состав, физико-химические свойства и механизм образования коацерватных капель. Установлено, что они обладают избирательной адсорбционной способностью по отношению к различным органическим веществам и способны включать в себя ферментные белки, катализирующие превращения находящихся в капле веществ. На примере распада крахмала до сахаров в коацерватной капле при участии фермента бета-амилазы было показано, что в коацерватах может происходить концентрированное образование продуктов реакции. В этом случае коацерваты правомочно рассматривать как системы, морфологическая структура которых существенно влияет на характер протекающего в них процесса.
Работами Т.Н. Евреиновой (1962) было установлено, что при образовании коацерватных капель происходит очень высокое концентрирование полимеров. Несмотря на свою жидкую консистенцию, капли обладают определенной внутренней структурой и резкой границей раздела с внешней средой. А.И. Опарин и К.Б. Серебровская (1963) показали, что при синтезе полиаденина в водном растворе гистона или полилизина можно наблюдать образование коацерватных капель, совершающееся при участии бактериальной полинуклеотидфосфорилазы. При этом равновесие реакции полимеризации резко смещается в сторону синтеза. Включив в коацерватные капли катализатор, эти исследователи получили модели систем, в которых протекают окислительно-восстановительные реакции, в том числе под влиянием света и хлорофилла.
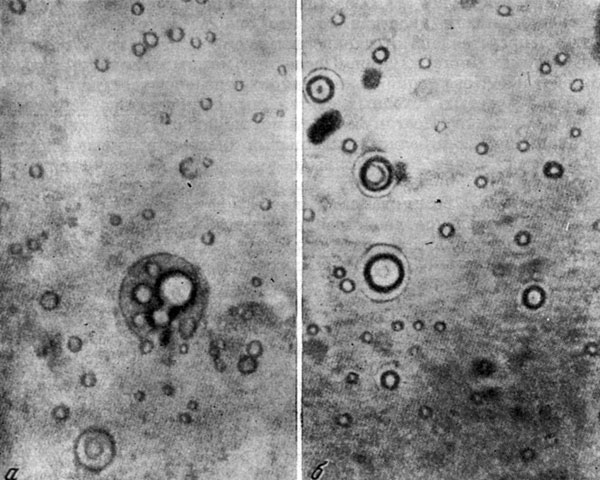
Коацерватные капли, содержащие полипептиды (а) и полинуклеотиды (б), по А.И. Опарину (1966).
Включение катализаторов приводило к неравенству концентрации веществ в капле и в среде и автоматически превращало капли в открытые микросистемы, характеризующиеся постоянным обменом веществ со средой. Это и явилось, вероятно, предпосылкой для возникновения «естественного отбора» таких систем, который способствовал сохранению наиболее устойчивых. Назвав условно такие системы «пробионтами», Опарин указывает, что в первую очередь должен был совершенствоваться их каталитический аппарат как главный фактор организации обмена веществ. Каталитическая активность простейших катализаторов была очень невелика, однако в ходе естественного отбора пробионтов создавались каталитические комплексы — коферменты, обладавшие повышенной активностью. Повсеместное распространение в живых организмах коферментов указывает на их очень древнее происхождение. При дальнейшем усложнении обмена коферменты перестали удовлетворять потребностям живых систем: прогрессивная эволюция пошла по пути образования множества новых, гораздо более мощных катализаторов — ферментов. Таким образом, согласно Опарину, происходило совершенствование как всей системы в целом, так и отдельных ее механизмов.
Многочисленные геологические исследования показали невозможность установления на земной поверхности термодинамического равновесия с неизбежным для него полным распадом абиогенно возникших органических веществ. Вместе с тем наряду с синтезом все более сложных органических веществ на одних и тех же территориях и акваториях мог иметь место и их распад, сопровождавшийся затем новым синтезом. Чередование таких процессов могло приводить к многократному возникновению пробионтов.
А.Г. Пасынский (1959) показал, что в открытых химических системах ферментативные реакции проявляют некоторые особенности, отсутствующие в замкнутых системах. По его мнению, наиболее существенное эволюционное значение должны были иметь открытые системы, устойчивость которых динамически поддерживалась происходившими в системе химическими реакциями. Другой важной особенностью открытых систем является то, что в условиях сложного комплекса сопряженных реакций основное значение приобретает направление, по которому реакция может протекать с наибольшей скоростью. По мнению Пасынского, на добиологической стадии развития принцип максимальной скорости реакции мог обеспечивать преимущество одних открытых систем перед другими и лежать в основе естественного отбора. По-видимому, наиболее примитивные формы живых существ могли образоваться на основе открытых химических систем еще до того, как возникли формы передачи информации, связанные с молекулами нуклеиновых кислот.
Общей чертой любой многомолекулярной системы, выделившейся иэ «первичного бульона», должно было быть наличие определенной поверхности, отделявшей эту систему от окружающего раствора. Такие поверхности возникают спонтанно в результате физико-химических закономерностей, заложенных в особенностях химического состава и структуры сложных органических и полимерных веществ. По мнению Г.А. Деборина (1967), появление фазовой границы с измененными структурно-механическими свойствами должно было неизбежно привести к локализации в ней ряда физических и химических процессов, связанных с переносом веществ из одной фазы в другую, а затем, в ходе дальнейшей эволюции, и некоторых биохимических процессов, сопутствующих как явлениям переноса, так и пространственного монтажа ферментных систем. Таким образом, можно полагать, что важнейшим структурным элементом первичных многомолекулярных систем явилась самоформирующаяся поверхностная пленка, которая в процессе дальнейшей химической эволюции могла подвергаться под действием отбора усложнению в составе и тонком строении. Было показано, что одновременно с общим процессом эволюции предбиологических систем путем естественного отбора происходило постепенное приспособление молекулярной поверхности раздела к выполнению определенных функций, их превращение в более упорядоченные, многомолекулярные структуры, специализированные на переносе веществ и энергии, необходимых для жизнедеятельности и размножения клетки. Эта эволюция достигла наибольшего совершенства в клеточных органеллах, например, в мембранах митохондрий, где сосредоточены ферментативные механизмы дыхания и окислительного фосфорилирования.
Микросферы С. Фокса.
В качестве одного из возможных предшественников клетки большой интерес представляют микросферы, впервые полученные С. Фоксом и сотрудниками (1955). Фокс считает, что белок мог впервые образоваться на поверхности Земли близ областей с повышенной температурой, где смесь накопившихся аминокислот нагревалась, полимеризовалась, а затем вымывалась в океан. Р. Янгу (1964, 1970) удалось показать, что в процессе синтеза протеиноидов из аминокислот образуются гуанин, а также жирные кислоты. Это делает микросферы интересным объектом для изучения одного из возможных путей первичного образования клеток.
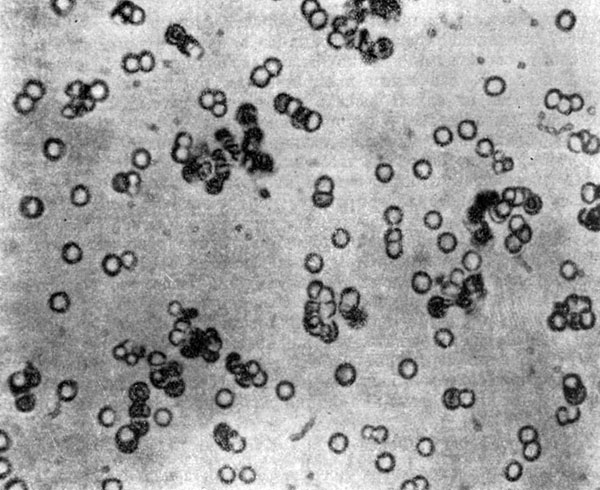
Микросферы, по С. Фоксу (1960).
* * *
Современная концепция возникновения жизни на Земле является результатом широкого синтеза биологических знаний, добытых исследователями разных специальностей. Особое значение для ее обоснования и развития имел прогресс биологической химии и переход к исследованиям живой материи на молекулярном уровне. Несмотря на то, что полностью воспроизвести цепь условий, которая привела однажды к возникновению жизни, по-видимому, никогда не удастся, современная наука уверенно восстанавливает все новые и новые звенья на пути от химической к биологической эволюции. Существенно отметить, что независимо от того, какой из путей образования индивидуальных многомолекулярных систем, исходных для дальнейшей эволюции, будет признан наиболее вероятным, общая теория возникновения жизни, по-видимому, останется незыблемой.
Теория возникновения жизни на Земле может лечь в основу более общей теории возможного возникновения жизни на других планетах, так как с несомненностью установлено, что синтез сложных органических соединений во Вселенной и процессы их химической эволюции носят более общий характер, чем это предполагалось первоначально. Таким образом, эта теория, помимо своего большого теоретического значения имеет и большой выход в практику, вооружая исследователей космоса научными подходами к поискам внеземных форм жизни.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК