Полиплоидные комплексы
Наиболее парадоксальные случаи видообразования путем гибридизации — так называемые полиплоидные комплексы, открытые и подробно изученные американским ботаником-эволюционистом Г. Ледьярдом Стеббинсом (род. 1906)[510]. Представим себе четыре вида А, В, С и D, каждый из которых имеет только ему свойственный хромосомный набор с соответствующими диплоидными числами, равными 2n(АА)=18, 2n(ВВ)=22, 2n(CС)=16, 2n(DD)=24. Каждый из этих видов репродуктивно изолирован! ото всех остальных, занимает свою экологическую нишу. Эти виды могут дать шесть аллотетраплоидных вариантов: Е (2n=40, геном ААВВ), F (2n=34, ААСС), G (2n=42, AADD), Н (2n=38, BBCC), I (2n=46, BBDD), J (2n=40, CCDD). Каждый из возникших аллотетраплоидов оказывается репродуктивно изолированным как от другого аллотетраплоида, так и от предковых видов A—D, т. е. генетически все они настоящие виды, а обладая различными генотипами и, соответственно, различными фенотипами, они смогут бороться за завоевание разных экологических ниш.
Если вновь, уже на уровне аллотетраплоидов произойдет нарушение процесса редукции числа хромосом в мейозе, то слияние тетраплоидных гамет может дать серию вариантов аллоокгоплоидов, например, AABBCCDD с 2n=80, т. е. форму, предками которой будут четыре диплоидных и два аллотетраплоидных вида. Такие формы, называемые полиплоидными комплексами, или компилоспециес (рис. 242), были получены в эксперименте и обнаружены в природе[511] у некоторых родов цветковых растений (ежи — Dactylus[512], хлопчатника — Gossypium, кларкии — Clarkia). Таким образом, предком одного вида, по крайней мере аллополиплоидного, может быть не один, а несколько видов.
То обстоятельство, что в ходе видообразования может происходить чередование дивергенции (кладогенеза) и процесса слияния разных ветвей одного ствола, было известно и равнее. Так, уже Ф. Г. Добржанский (1937)[513] отчетливо представлял, что далеко не всегда новый вид образуется из одной популяции. Такую эволюцию Добржанский назвал ретикулярной, или сетчатой эволюцией (рис. 243). Однако когда говорят о ретикулярной эволюции, то обычно имеют в виду слияние генофондов нескольких популяций одного вида или же подвидов внутри вида, а начиная с видового уровня, когда возникает репродуктивная изоляция, говорят лишь о дивергентной эволюции. Истинная картина, наблюдающаяся в природе, как мы видели, может быть гораздо более сложной. Слияние генофондов разных видов в случае растений и животных с наружным оплодотворением может идти лишь за счет нарушений мейоза без разрушения иных эволюционных барьеров.
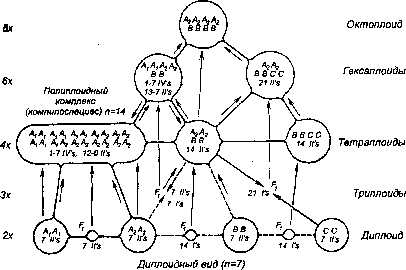
Рис. 242. Схема происхождения компилоспециеса (полиплоидного комплекса), родоначальниками которого являются несколько «старых» видов.
По Л. Стеббинсу (1977) из Н. Н. Воронцова (1980).
Аллополиплоидия[514], возникновение полиплоидных комплексов[515] — все это примеры макроэволюции, идущей своеобразным путем. Здесь неуместно говорить о том, что микроэволюционные процессы предшествовали видообразованию (хотя не подлежит сомнению, что микроэволюционные процессы изменяли генотипические составы популяций каждого из предковых видов, что выживание возникших аллополиплоидов во многих случаях будет зависеть от таких микроэволюционных событий, как возникновение точковых мутаций отдельных генов, увеличение частот аллелей, повышающих приспособленность полиплоидов к каким-то конкретным условиям среды, и т. п.).

Рис. 243. Ретикулярная эволюция.
Из: Th. Dobzhansky (1951).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК