Формы видообразования во времени
Дарвиновская схема дивергенции на самом деле демонстрировала не только дивергентную эволюцию, но и эволюцию без распадения исходной ветви на дочерние. Возможность дивергентного происхождения таксонов от одной предковой популяции ныне никем не отрицается. Б. Рент[486] в 1954 г. и Дж. Хаксли[487] в 1957 г. предложили специальные термины для трех форм видообразования во времени (рис. 235), выделенных еще Дарвином.
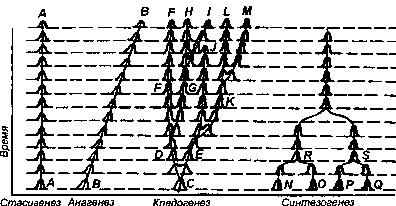
Рис. 235. Четыре основные формы видообразования во времени. Первые три формы видообразования были ясны еще Дарвину. Синтезогенез (симгенез) выделен автором В 1980 Г.
Из Н. Н. Воронцова (1984).
Анагенез — процесс прогрессивного развития группы, не связанный с распадением ее на боковые ветви. Эволюция предков человека от австралопитеков через человека умелого (Homo habilis) и человека прямоходящего (Homo erectus) к человеку разумному была, в основном, анагенетическим процессом. Анагенез связан с действием движущей, или ведущей, формы естественного отбора.
Кладогенез — процесс увеличения числа ветвей в группе. Только к кладогенезу и относятся понятия дивергентной эволюции[488] и адаптивной радиации. Многочисленные примеры кладогенезов, связанные с идиоадаптациями А. Н. Северцова, дают группы, эволюировавшие в условиях ненасыщенных биогеоценозов изолированных территорий. Дивергенция австралийских сумчатых, давших на этом континенте сумчатого волока, сумчатую куницу, сумчатую летягу, сумчатых мышей и сумчатого крота, — наиболее популярный пример кладогенеза. Однако справедливости ради следует отметить, что все приведенные виды относятся не только к разным видам (и родам), но и к разным семействам отряда сумчатых.
Как это ни странно, конкретных примеров кладогенезов на уровне видов одного рода, для которых был бы изучен палеонтологический материал, крайне мало. В таких случаях часто приводят примеры дивергенции видов одного рода на разных островах одного архипелага. К числу таких косвенных свидетельств кладогенеза относятся знаменитые дарвиновы вьюрки Галапагосских островов[489] (рис. 236), гавайские цветочницы[490] (рис. 237), моллюски акантеллиды тех же Гавайских островов и иные аналогичные примеры. В результате у читателей создается впечатление, что дивергентная эволюция идет только лишь где-нибудь на Соломоновых островах. Конечно, это не так, однако почти полное отсутствие хорошо документированных случаев дивергентной эволюции на уровне видов одного рода говорит о том, сколь мало мы знаем даже о наиболее обычной форме видообразования. Кладогенез связан с одновременным или последовательным действием не только движущей, но и дизруптивной (разрывающей) формы отбора.
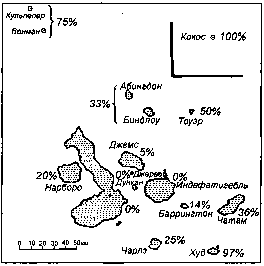
Рис. 236. Увеличение процента эндемичных видов дарвиновых вьюрков (Geospiza) на Галапагосских островах по мере движения к периферии архипелага как косвенное свидетельство дивергентной эволюции в условиях изоляции.
Из: D. Lack (1947).
Дальнейший анализ, в том числе и эксперименты по моделированию дивергентной эволюции на ЭВМ[491], свидетельствуют, что дивергентная эволюция теоретически возможна и без территориальной изоляции[492]. Поскольку в условиях изоляции интенсивно протекают генетикоавтоматические процессы, мы должны признать, что эти процессы могут приводить к кладогенезу и при отсутствии естественного отбора.
Стасигенез — процесс длительного сохранения вида или иного таксона без прогрессивного изменения в каком-либо направлении, без направленного отбором исторического развития в каком-то направлении (т. е. без анагенеза) и без дивергенции (без кладогенеза). По пути стасигенеза развиваются немногие переживающие свой век представители предковых групп. Реликтовые ракообразные — мечехвосты, кистеперая рыба — латимерия, реликтовое плеченогое — лингула, примитивная рептилия — гатгерия, сохранившаяся лишь на немногих островках у берегов Новой Зеландии — вот примеры таксонов, развивавшихся в последний период своей эволюции путем стасигенеза.
При стасигенезе в течение многих тысяч поколений действует стабилизирующая форма отбора, сохраняющая на протяжении тысячелетий то же среднее значение данного признака. Обнаружение подобных реликтов, подчас неотличимых от своих предков, живших сотни миллионов лет назад, нередко вызывает появление в широкой печати статей об остановке эволюции, о существований видов, не эволюировавших сотни миллионов лет, и т. п. На самом деле проявление стабилизирующей формы отбора не есть отсутствие отбора; отбор на поддержание одной и той же средней не есть отсутствие эволюции. При снятии пресса стабилизирующей формы отбора вариационная кривая изменчивости вида начинает расплываться, в популяции накапливаются такие мутации, какие при стасигенезе неминуемо отсекаются стабилизирующей формой отбора. Этот процесс дестабилизации онтогенеза и расширения диапазона изменчивости при снятии стабилизирующей функции отбора был тщательно изучен в известных опытах акад. Д. К. Беляева (1917—1985, рис. 238) и его ученицы Л. Н. Трут по изучению изменчивости у лисиц в процессе их приручения.
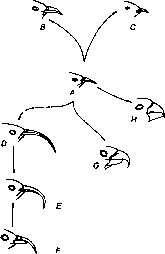
Рис. 237. Дивергенция по форме клюва и кладогенез гаванских цветочниц (Drepanididae).
По Е. О. Dodson из Ст. Скаврона (1965).
Как показали эти исследования, в линии лисиц, селекционированных на дикое поведение, изменчивость оставалась небольшой, в то время как в линии лисиц, отбиравшихся по принципу приручаемости, наблюдалось такое же резкое расширение диапазона изменчивости, какое отличает настоящих домашних зверей от их диких сородичей. Такое расширение спектра изменчивости при доместикации Д. К. Беляев связывал с дестабилизирующим отбором, что представляется мне неверным. Дестабилизация онтогенеза, вызванная снятием пресса стабилизирующего отбора, и дестабилизирующий отбор — это разные явления. Любое снятие стабилизирующего отбора ведет к неминуемому расширению диапазона изменчивости популяции как за счет выщепления рецессивных мутаций, так и за счет появления новых селективно нейтральных мутаций. При дестабилизирующем[493] (или центростремительном) отборе идет отбор новых мутаций с тем же значением средних величин, но с более широкой нормой реакции генотипа.
К. М. Завадский[494] предложил называть видообразование путем ветвления (т. е. кладогенез) сегрегациогенезом, а видообразование путем слияния разнородных таксонов именовать синтезогенезом. На самом деле в природе существует не две, а четыре формы видообразования.
Сейчас становится все более очевидным, что в природе, наряду с кладо-, ана- и стасигенезом, существует и достаточно широко распространена еще одна форма происхождения новых таксонов — путем аккумуляции геномов разных, ранее репродуктивно изолированных ветвей. Такую форму эволюции я предложил назвать симгенезом. Этот термин стал входить в литературу[495]. Вскоре после публикаций, в которых мною обосновывалась роль симгенеза в эволюции[496], вышла интересная работа ленинградских герпетологов Л. Я. Боркина и И. С. Даревского[497], где . приведены убедительные примеры гибридогенного происхождения видов у позвоночных животных. Новейшие исследования В. П. Васильева[498] показали большую роль полиплоидии и гибридогенеза в видообразовании рыб. Е. Н. Панов[499] успешно исследует роль гибридизации в формообразовании у птиц. Работы нашей лаборатории говорят о том, что на стыках ареалов аллопатрических форм, в частности, у сусликов (Spermophilus) или сурков (Marmota), существуют зоны диффузии генов через межвидовые изоляционные барьеры. Еще совсем недавно считалось, что гибридогенное видообразование наблюдается лишь в отдельных группах растений.
В чем состоит принципиальное отличие симгенеза (синтезогенеза) от сегрегациогенеза? При анагенезе и кладогенезе процесс видообразования может носить (но не обязательно) постепенный, градуалисгический характер. В этом случае можно говорить о том, что налэпление отдельных мутаций структурных генов, обеспечиваемое как отбором, так и генетико-автоматическими процессами (а нередко и совместным действием этих двух формирующих факторов эволюции), может идти постепенно. Так же постепенно (нередко вообще как побочный эффект дивергенции по другим признакам) формируются барьеры изолирующих механизмов эволюции, которые со временем могут стать абсолютными. Таким образом, в случае анагенеза и кладогенеза макроэволюция действительно может идти через микроэволюцию в соответствии с одним из постулатов СТЭ.
Иначе обстоит дело при симгенезе. Даже гибридогенное видообразование, не говоря уже о случаях образования таксонов более высокого ранга путем симгенеза, не может быть сведено к классическим микроэволюционным процессам, здесь нет элементарного эволюционного явления — изменения генотипического состава популяции, ибо гибридная популяция ранее репродуктивно изолированных видов есть нечто принципиально новое: если в случае анагенеза и кладогенеза отбор, изоляция и генетико-автоматические процессы вызывают и направляют эволюцию, то в случае синтезогенеза инициирующим фактором является объединение разных геномов.
Бесспорно, что это объединение разных геномов и создание нового сбалансированного генома идет на фоне действия естественного отбора, отбрасывающего нежизнеспособные комбинации геномов. Но как элиминируемые, так и отбираемые индивидуумы в случае гибридогенеза оказываются изначально репродуктивно изолированными от родительских видов. Таким образом, синтезогенез должен рассматриваться (в случае возникновения таксона ранга вида и выше за счет объединения геномов разных видов) как макроэволюционный процесс, не вызванный микроэволюционными процессами. Вместе с тем ясно, что вслед за возникновением макроэволюционного акта начнут действовать на уровне новой генетической системы микро-эволюционные процессы.
Наиболее понятна и абсолютно доказана роль симгенеза на низших этапах макроэволюции, на уровне образования таксонов ранга вида, рода. Вместе с тем накапливаются данные, говорящие о том, что и мегаэволюция может идти за счет процессов интеграции разнородных геномов. Мы рассмотрим эти два уровня интеграции геномов порознь.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК