История синтеза генетики и дарвинизма
В 1905 году ученик М. А. Мензбира и Н. К. Кольцова (рис. 181) по Московскому университету зоолог Сергей Сергеевич Четвериков (1880—1959, рис. 182) обратил внимание на то обстоятельство, что периодические колебания численности (или «волны жизни» по Четверикову) могут влиять на направление и интенсивность давления естественного отбора[352]. Позднее стало ясно, что пульсации численности вида не могут быть безразличны для генотипического состава популяций. В годы депрессии численности преимущественно вымирают одни генотипы, а в годы последующих пиков численности генотипический состав новых временных популяций (или хронопопуляций) с неизбежностью имеет иное частотное распределение аллелей (взаимоисключающих состояний одного и того же гена), чем в предыдущую волну.

Рис. 181. Николай Константинович Кольцов.
Из: В. Полынин, 1969.

Рис. 182. Сергей Сергеевич, Четвериков.
Фото из архива автора, 1969.
В 1921 г. в Советскую Россию приехал ближайший сотрудник Моргана, будущий лауреат Нобелевской премии Герман Мёллер (1890—1967, рис. 183), рассказавший Кольцову и Четверикову о работах школы Моргана по хромосомной теории наследственности, выполненных на разводимых в пробирках американских видах дрозофил. Энтомологу Четверикову стало ясно, что с не меньшим успехом можно работать и на европейских видах этого рода. В отличие от американских генетиков, Четвериков решил изучать природные популяции дрозофил, личинки которых питаются грибами[353].

Рис. 183. Герман (Германович) Мёллер (второй слева) — сотрудник Н. И. Вавилова, член-корреспондент АН СССР — и его ассистент К. Офферман (третий слева) с советскими генетиками академиком АН УССР И. И. Аголом (крайний слева) и основателем Медико-генетического института С. Г. Левитом. И. И. Агол и С. Г. Левит были расстреляны. Мёллер в 1937 г. уехал из СССР в республиканскую Испанию, а затем в США. Фото начала 1930-х годов.
Из: Baumunk und Rie? (1994).
Активная творческая деятельность Четверикова в Кольцовском институте была, увы, непродолжительной. «Этот период охватывает короткие 8 лет с 1921 по 1929 г., но он очень насыщен и оставил глубокий след в истории отечественной и мировой генетики. Именно тогда под руководством Сергея Сергеевича сформировался очень активный коллектив исследователей, одна из сильнейших школ, положивших начало экспериментальной генетике и, в частности, эволюционной генетике в нашей стране»[354]. Четвериков организовал на Звенигородской биостанции и в кольцовском Институте экспериментальной биологии (ныне — Институт биологии развития им. Н. К. Кольцова РАН) широкие исследования по поиску мутаций в природных популяциях, а также знаменитый семинар «Дрозсоор» («совместный ор о дрозофиле»), вокруг которого зародилась отечественная, а вслед за ней и мировая генетика природных популяций[355].
За свою долгую и трудную жизнь Четвериков опубликовал лишь 33 работы, из которых статья 1926 г. «О некоторых моментах эволюционного процесса с точки зрения современной генетики» обессмертила его имя. В этой работе Четвериков показал, что:
1) мутационный процесс происходит в природных популяциях (что тогда было совсем не очевидно);
2) большинство вновь возникших мутаций понижает жизнеспособность, хотя изредка возникают и мутации, повышающие таковую;
3) в условиях свободного скрещивания вид достаточно устойчив и сохраняет (в соответствии с законом Харди— Вайнберга) исходное соотношение частот аллелей, а каждая рецессивная мутация «впитывается видом в гетерозиготном состоянии» и при отсутствии отбора может сохраняться в том же соотношении неограниченно долго;
4) гетерозиготность по разным аллелям разных генов «пропитывает вид во всех направлениях» и в результате случайных комбинаций постепенно «заражает» большинство индивидов вида;
5) по мере старения вида в нем накапливается все большее число мутаций, при этом признаки вида расшатываются;
6) генетическая изменчивость проявляется наиболее сильно, когда многочисленный вид распадается на ряд небольших, изолированных колоний;
7) наиболее обычна пространственная изоляция, хотя может существовать и временная экологическая изоляция [Здесь можно усмотреть развитие идей немецкого зоолога и путешественника Морица Вагнера (1813—1887), выпустившего в 1868 г. книжку «Теория Дарвина и закон миграции организмов»[356], впервые высказавшего мысль о том, что один отбор без пространственной изоляции не может привести к возникновению нового вида. Эти идеи были обобщены в посмертно изданном сборнике его избранных работ «Происхождение видов путем пространственной изоляции» (1889). От Четверикова представления о роли изоляции в видообразовании воспринял Н. В. Тимофеев-Ресовский, выдвинувший в конце 30-х гг. изоляцию как один из элементарных эволюционных факторов. В 40-е и последующие годы идеи М. Вагнера уже на новом популяционном уровне знаний активно пропагандировал и пропагандирует Эрнст Майр];
8) изоляция, наряду с наследственной изменчивостью, «является основным фактором внутривидовой (а следовательно, и межвидовой) дифференциации» [Из этого высказывания Четверикова возникнет затем тезис Тимофеева-Ресовского и многих других создателей СТЭ о том, что макроэволюцш идет только через микроэволюцию. Существенно то, что Четвериков уже в 1926 г. называет изоляцию основным фактором эволюции!];
9) естественный отбор является антагонистом свободного скрещивания — панмиксии;
10) приспособительная эволюция без изоляции ведет к полной трансформации вида, но не может привести к его распадению на два вида [Важнейшее заключение! Четвериков представлял возможность недивергентного, анагенетического (см. ниже) видообразования без изоляции и считал, что дивергентное видообразование, или кладогенез, невозможно только на основе отбора без изоляции];
11) отбор, как и свободное скрещивание, ведет к накоплению рецессивных аллелей в гетерозиготном состоянии;
12) свободное скрещивание — панмиксия — ведет к повышению полиморфизма вида, а отбор ведет к мономорфизму;
13) отбор отбирает не только отдельный ген, контролирующий отдельный признак, но и, благодаря множественности (плейотропносги) действия генов, влияет на все генное окружение отбираемого гена, на его генотипическую среду.
В этой статье, равно как и в более ранних выступлениях и в работах, сделанных с учениками, С. С. Четвериков впервые показал пути использования генетических и эволюционных подходов для изучения изменчивости в природных популяциях. Ученики Четверикова по кружку — Николай Владимирович Тимофеев-Ресовский (1900—1981), Елена Александровна Тимофеева-Ресовская (1899—1973), Николай Константинович Беляев (1899—1937), Борис Львович Acmaypoe (1904—1974), Елизавета Ивановна Балкашина (1899—1981), Дмитрий Дмитриевич Ромашов (1899—1963), Александр Николаевич Промптов (1898—1948), Петр Фомич Рокицкий (1903—1977), Сергей Романович Царапкин (1892—1960), Сергей Михайлович Гершензон (1906—1998); последователи Четверикова — Феодосий Григорьевич Добржанский (1900—1975), ученик ленинградского генетика, зоолога и эволюциониста Юрия Александровича Филипченко (1882—1930); Николай Петрович Дубинин (1907—1998), ученик московского генетика и эволюциониста Александра Сергеевича Серебровского (1892—1948) и сотрудник Кольцова — заложили в конце 20—начале 30-х гг. основы генетики природных популяций (рис. 184—195). И хотя Четвериков в 1929 г. был арестован и, несмотря на хлопоты Н. К. Кольцова, К. С. Станиславского и наркома Н. А. Семашко, выслан на Урал и во Владимир, прекратил свои популяционные работы и навсегда покинул Москву, а «Дрозсоор» распался и от лаборатории остались «рожки да ножки», позже она все же возродилась на новой основе под руководством Н. П. Дубинина[357].

Рис. 184. Николай Владимиович Тимофеев-Ресовский. Фото Esko Suomalainen (1943).
Из архива автора.
Рис. 185. Елена Александровна Тимофеева-Ресовская (урожденная Фидлер).
Из: Н. В. Тимофеев-Ресовский (1995).

Рис. 186. Николай Константинович Беляев.
Из архива Н. Б. Астауровой.

Рис. 187.Борис Львович Астауров.
Фото С. И. Раджабли.
Судьбы участников «Дрозсоора» типичны для судеб отечественной интеллигенции тех лет[358]. Н. К. Беляев уехал после ареста Четверикова в Тбилиси, занимался генетикой шелкопряда; был арестован и расстрелян в 1937 г. Б. Л. Астауров уехал в Среднюю Азию, выжил, стал академиком АН СССР, в 1966 г. воссоздал Кольцовский институт (разогнанный в 1948 г.) и был его директором. Автор классических работ по регуляции пола у шелкопряда. Е. И. Балкашина вместе с сестрой отправилась в ссылку в Восточный Казахстан, работала энтомологом на Усть-Каменогорской санэпидстанции и отошла от науки. Ее совместные с Б. Л. Астауровым исследования макромутаций широко цитировались Р. Гольдшмидтом (1940) в его монографии о макроэволюции. Д. Д. Ромашов под влиянием стресса на несколько лет попал в психиатрическую клинику, выжил и стал одним из ведущих теоретиков популяционной генетики; с 1948 по 1955 г. был безработным.

Рис. 189. Дмитрий Дмитриевич Ромашов.
Из архива Н. Б. Астауровой.

Рис. 188. Елизавета Ивановна Балкашина.
Из архива автора.

Рис. 190. Александр Николаевич Промптов.
Из: А. Н. Промптов (1956).

Рис. 191. Петр Фомич Рокицкий.
Из: Гершензон (1992).
А. Н. Промптов уехал в 1937 г. в Нижний Новгород, затем в Колтуши под Ленинградом, прославился как натуралист своей книгой «Птицы в природе», занимался генетикой поведения животных. После августовской сессии ВАСХНИЛ 1948 г. покончил с собой. П. Ф. Рокицкий был арестован, позднее работал в области генетики домашних животных под Москвой, после 1948 г. оказался в Коми филиале АН СССР, занимался биометрией и статистикой. После 1965 г. получил хорошие возможности в Минске, стал академиком АН Белоруссии. Н. В. и Е. А. Тимофеевы-Ресовские и С. Р. Царапкин в 1925—1945 гг. работали в Германии и, казалось бы, избежали участи своих московских товарищей по семинару. Но в 1945 г. Н. В. Тимофеев-Ресовский и С. Р. Царапкин были арестованы, получили по 10 лет заключения с последующим «поражением в правах», были посмертно реабилитированы лишь в конце 1991 г. Н. В. Тимофеев-Ресовский был избран членом многих академий мира, но не у себя на родине. С. М. Гершензон, сын крупнейшего культуролога России М. О. Гершензона, автор классических работ в области популяционной генетики 30—40-х гг., открыл мутагенное действие ДНК в 40-х гг. и вплотную подошел к пониманию роли ДНК в явлениях наследственности. Ему принадлежат первые работы по предсказанию обратной передачи информации с РНК на ДНК. В 1948 г. эти исследования были запрещены, а Нобелевские премии достались зарубежным ученым. С. М. Гершензон стал академиком АН Украины. Так сложились судьбы четвериковской школы.

Рис. 192. Сергей Михайлович Гершензон.
Из: Гершензон (1992).
В начале 1930-х гг. Н. К. Кольцов пригласил в свой институт Н. П. Дубинина — ученика А. С. Серебровского и Д. Д. Ромашова. «Через Д. Д. Ромашова была передана эстафета эволюционной генетики, и факел, зажженный ранее четвериковской школой, — писал Б. Л. Астауров, — не погас. В дальнейших исследованиях Тимофеевых-Реовских за рубежом, лаборатории Н. П. Дубинина в Кольцовском институте, многих других советских генетиков (особенно А. А. Малиновского, P. Л. Берг, В. С. Кирпичникова, Ю. М. Оленова,) и в лаборатории Ф. Г. Добржанского за рубежом линия эволюционной и популяционной генетики, начатая в лаборатории С. С. Четверикова, получила дальнейшее развитие»[359].

Рис 193. Юрий Александрович Филиппенко. Основатель петроградско-ленинградскои школы генетиков. Эволюционист. Ввел понятия «микроэволюция» и «макроэволюция». Созданная им лаборатория генетики АН СССР после его кончины возглавлялась Н. И. Вавиловым, который преобразовал ее в Институт генетики АН СССР (ныне — Институт общей генетики им. Н. И. Вавилова РАН).
Из: Развитие биологии в СССР (1967).
Действительно, несмотря на то, что работа Четверикова была опубликована по-русски, а публикации его учеников в отечественных журналах вышли с большим опозданием и в неполном виде, его идеи были быстро разнесены по свету. Н. В. Тимофеев-Ресовский неустанно пропагандировал идеи своего учителя среди европейских генетиков, а Ф. Г. Добржанский — среди американских. Следует также отметить, что оба эти исследователя были по своему изначальному образованию зоологами, и потому они смогли пропагандировать идеи Четверикова не только в генетических, но и в зоологических кругах.

Рис. 194. Феодосий Григорьевич Добржанский.
Из: Dobzhansky’s Genetics of Naturals Populations (1981).
Обмену многими идеями и методами между ведущими тогда генетическими школами — американской и советской — способствовало несколько обстоятельств: многие работы советских генетиков печатались на европейских языках, в 20-е годы многие советские генетики выезжали за рубеж на длительные сроки (рис. 196—198). Среди них были Н. И. Вавилов, Н. К. Кольцов, блестящий ученик Вавилова безвременно погибший в сталинских тюрьмах цитогенетик Георгий Дмитриевич Карпеченко (1899—1942), классик цитогенетики растений Михаил Сергеевич Навашин (1896—1973), генетик и селекционер растений Антон Романович Жебрак (1901—1965), а в конце 20 — начале 30-х гг. в СССР по приглашению Вавилова долгое время работали ученики Моргана — Мёллер (1890—1967), которого советские коллеги звали Германом Германовичем, будущий Нобелевский лауреат, избранный в те годы членом-корреспондентом АН СССР; Кэльвин Бриджес (1889-1938) — один из создателей хромосомной теории наследственности, а также ведущий специалист по генетике хлопчатника С. Е. Харланд.

Рис. 195. Александр Ссргеенич Серебровский (слева) — создатель школы генетиков в МГУ; руководитель работ по теории дробимости гена. Ввел понятия «геногеография» и «феногеография». Предложил генетические методы регуляции численности насекомых. Николай Петрович Дубинин (справа). Москва, зоотехнический ин-т, 1928 г.
Из архива автора.
Крупнейший английский генетик-эволюционист Джон Бердон Сандерсон Холдейн (1892-1964, рис. 199) встречался с Четвериковым на V Международном генетическом конгрессе в Берлине в 1927 г., в его лаборатории был осуществлен перевод четвериковской статьи 1926 г., в работах его ученицы Сесиль Гордон (1936, 1939) отмечалась инициирующая роль работы Четверикова в последующих исследованиях; этим переводом пользовались сотрудники и гости лаборатории Холдейна.
Наряду с развитием генетики природных популяций, которое было инициировано Четвериковым, возникает и теоретическая популяционная генетика. Классические работы англичанина Рональда Фишера (1890—1962, рис. 200) — математика и теоретического физика по образованию, исследовавшего теорию доминантности, вероятностные аспекты теории мимикрии, выпустившего в 1930 г. книгу «Генетическая теория естественного отбора»[360], американского теоретика Сьюэлла Райта (1889—1988, рис. 201) — зоолога по образованию и генетика животных по опыту работы, в 1930 г. откликнувшегося обширной рецензией на книгу Р. Фишера, а в 1931 г. выпустившего монографию «Эволюция в менделевских популяциях»[361], независимо от Четверикова, хотя и с опосредованным участием его школы, положили начало синтезу генетики и дарвинизма на Западе.

Рис. 196. Советские генетики (все в шляпах) на экскурсии по Берлину во время Международного генетического конгресса (1927). Слева направо: Ю. А. Филипченко, 11. И. Вавилов, Г. А. Левитский, С. С. Четвериков, Н. В. Тимофеев-Ресовский, Г. Д. Карпеченко (сидит). Последний массовый выезд отечественных генетиков за границу перед закрытием «железного занавеса».
Из: В.-М. Baumunk und J. Rie? (1994).

Рис. 197. Интернациональный коллектив учеников Т. Г. Моргана (в центре) в Калифорнийском институте технологии («КалТек»), Пасадена, 1930 г. Во втором ряду — Рокфеллеровские стипендиаты Ф. Г. Добржанский (второй слева), Г. Д. Карпеченко (третий слева). Пятый слева — К. Бриджес.
Из: В.-М. Baumunk und J. Rie? (1994).
Рис. 198. Н. И. Вавилов, Т. Г. Морган и Н. В. Тимофеев-Ресовский на Международном генетическом конгрессе в Итаке (США, 1932). Н. И. Вавилов — вице-президент конгресса — был единственным приехавшим из СССР. Н. В. Тимофеев-Ресовский прибыл из Берлина.
Здесь следует обратить внимание на одно важное заключение из работы Четверикова (1926): «У нас нет оснований отрицать возможность неадаптивной эволюции. Во многих случаях можно предполагать, что существующие адаптивные различия между близкими формами были не причиной расхождения последних, а, напротив, специфический характер этих адаптивных признаков является следствием уже ранее наступившего обособления форм»[362]. Эта идея была развита Р. Фишером, С. Райтом, Н. П. Дубининым, Д. Д. Ромашовым, а затем, на основе новейших открытий молекулярной биологии, была вновь подхвачена в 60-х гг. М. Кимурой[363], Дж. Кингом[364] и другими сторонниками так называемой «недарвиновской эволюции».

Рис. 199. Джон Б. С. Холдейн.
Из: Genetics (1965).

Рис. 200. Рональд Эйлмер Фишер.
Из: R. W. Clark (1984).

Рис. 201. Сьюэлл Райт.
Из: Dobzhansky’s Genetics of Naturals Populations (1981).
Фишер и Райт обратили внимание на тот важный факт, что распределение и концентрация частот аллелей в популяциях может идти не только под действием панмиксии (неограниченного перемешивания) и естественного отбора, но и под действием случайных, стохастических факторов, получивших на Западе название «дрейфа генов».
Предложенное в 1908—1909 гг. английским математиком Годфреем Гарольдом Харди (1877—1947) в статье «Менделевские соотношения в смешанной популяции»[365] и немецким врачом Вильгельмом Вайнбергом (1862—1937) в статье «О закономерностях наследственности у человека»[366] правило Харди—Вайнберга, объясняющее сколь угодно долгое сохранение в популяции двух аллелей одного гена (доминантного А и рецессивного а), действует лишь при следующих ограничениях: 1) бесконечно большая популяция; 2) абсолютная панмиксия — неограниченность вариантов скрещивания; 3) отсутствие мутационного процесса; 4) отсутствие различий в селективной ценности аллелей; 5) отсутствие перекрывания поколений во времени (что свойственно, например, многим лососевым рыбам, цикадам, майским хрущам, но не свойственно подавляющему большинству других групп).
Несоблюдение этих идеальных условий приводит, даже в случае одинаковой селективной ценности аллелей А и а, к случайному вытеснению одного аллеля другим, так что в конечном счете одни популяции могут со временем оказаться мономорфными по аллелю А, а другие по аллелю а. Это случайное вытеснение одного селекционно нейтрального (т. е. с равновероятным выживанием под действием сходного естественного отбора) аллеля другим и было названо «дрейфом генов».
Дальнейшее развитие теоретическая популяционная генетика получила в трудах Д. Д. Ромашова[367], Н. П. Дубинина[368], Тимофеева-Ресовского[369], Дж. Холдейна[370], А. А. Малиновского, С. Ли[371] . В 1935 г. в СССР Андрей Николаевич Колмогоров (1903—1987, рис. 202) математически показал, что наибольшие темпы изменения генотипического состава популяции характерны не для больших и не для малых популяции, а для серии полуизолятов с периодически возникающими и исчезающими потоками генов между ними [372].

Рис. 202. Андрей Николаевич Колмогоров.
Фото 1930-х гг. из архива автора.
В отличие от Фишера и Райта[373], исследовавших в 1930—1931 гг. динамику концентрации аллелей в популяциях при свободном скрещивании, Колмогоров рассмотрел уклонения от формулы Харди—Вайнберга в условиях частичной изоляции. Колмогоров подтвердил справедливость выдвинутого Малиновским представления «о существовании оптимума частичной изоляции для отбора рецессивных генов»[374].
Ленинградский генетик Юрий Александрович Филипченко (1882—1930) в 1927 г. предложил разделить эволюцию на два процесса — микроэволюцию, то есть дивергенцию от популяционного до видового уровня, и макроэволюцию, то есть дивергенцию на уровне выше вида[375]. По мнению Филипченко (в этом отношении он резко расходился как с рядом современных ему, так и с большинством последующих эволюционистов), роды произошли иным путем, чем виды. Он считал, что мутации, их комбинации и отбор могут объяснить лишь микроэволюцию[376]. Любопытно, что сейчас многие критики синтетической теории эволюции возвращаются к подобной позиции.
Термины Ю. А. Филипченко «микроэволюция» и «макроэволюция» широко пропагандировались на Западе Добржанским и Тимофеевым-Ресовским. В отличие от Филипченко, эти ученые считали, что макроэволюционный процесс и его закономерности сводимы к микроэволюционным процессам и их закономерностям.
В конце 1927 г. Ф. Г. Добржанский стал сотрудником Т. Г. Моргана, весьма критически относившегося к возможности синтеза генетики и дарвинизма[377]. Морган неоднократно подчеркивал пропасть между «натуралистами» и «учеными»: Добржанский, как и Четвериков, и как Тимофеев-Ресовский, не укладывался в построенную Морганом схему — он начинал как энтомолог-натуралист. Морган считал мутации аномалиями, полученными в эксперименте и не встречающимися в природе. Когда Морган узнал о работе Четверикова, о работах Тимофеева-Ресовского и Дубинина, продолживших это направление, он не проявил к ним того интереса, который проявили его ученики Г. Мёллер, А. Стертевант и другие.
В 1937 г. Добржанский выпустил небольшую по объему книгу «Генетика и происхождение видов»[378], в которой показал возможность приложения популяционногенетических данных к решению проблемы видообразования. Он сформулировал в этой книге продуктивное представление о системе «изолирующих механизмов эволюции» — тех барьеров, которые обеспечивают, с одной стороны, обособленность одного вида от другого, а с другой — способствуют поддержанию генетического единства вида как целого. В этой же книге было введено в широкий научных обиход полузабытое к тому времени правило Харди— Вайнберга.
В 1939—1943 гг. Тимофеев-Ресовский[379] предложил выделять элементарные эволюционные акты: элементарный эволюционный материал (мутация), элементарное эволюционное явление (изменение генотипического состава популяции), элементарную единицу эволюции (популяция). В этом стремлении выделить элементарные, далее не членимые акты в эволюционном процессе видно несомненное влияние длительного общения Н В. Тимофеева-Ресовского с физиками школы Н. Бора, проходившего в течение 30-х гг. При несомненном сходстве популяционно-генетических работ Добржанского и Тимофеева-Ресовского, при сходстве объектов (дрозофилы, «божьи коровки»), между эволюционными работами этих исследователей, развивавших традиции четвериковской школы, были и отличия. Добржанский, в отличие от Тимофеева-Ресовского, стал широко использовать для популяционных и сравнительных исследований изучение строения гигантских политенных хромосом личинок дрозофил. По этому же пути пошел и Н. П. Дубинин, успешно осуществивший вместе с Б. Н. Сидоровым, Н. Н. Соколовым, Б. Ф. Кожевниковым, Г. Г. Тиняковым ряд классических исследований цитогенетических особенностей природных популяций дрозофил[380]. Наибольший интерес в эволюционном отношении представляли работы Н. П. Дубинина и Б. Ф. Кожевникова, в которых целенаправленно были получены из обычных 8-хромосомных дрозофил 6- и 10-хромосомные формы, репродуктивно изолированные от родительского вида. Аналогичная работа на растениях была выполнена Е. Н. Герасимовой-Навашиной[381] на растении скерда (Crepis): ею был получен совершенно новый кариотип, в котором растение не содержало ни одной не перестроенной хромосомы предкового вида (рис. 203). Эти работы, выполненные задолго до появления модных слов о «генной и клеточной инженерии», были, по сути дела, одними из первых попыток конструирования генома с заранее спроектированными свойствами.
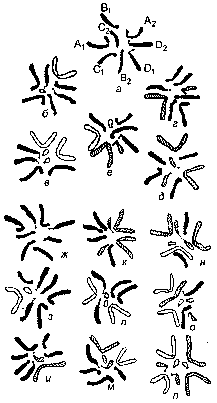
Рис. 203. Схема получения кариотипа нового синтезированного вида Crepis nova:
а — хромосомы Cr. tectorum; б, г — исходные гетерозиготные транслоканты; в, д — те же транслокацни, но переведенные в гомозиготное состояние; г — гибрид между в и д; ж-п — девять возможных комбинаций хромосом в потомстве е; п — Cr. nova.
Е. Н. Герасимовой из: Н. Н. Воронцов.
Идеи Четверикова роли явления рецессивности в эволюции с помощью Н. В. Тимофеева-Ресовского и Ф. Г. Добржанского распространились по мировому сообществу генетиков и эволюционистов и стали восприниматься как самоочевидные. В нашей стране на эти идеи особое внимание обратил Иван Иванович Шмальгаузен (1884—1963). В самом деле, ведь отбор идет по фенотипам, и, таким образом, особи с генотипами АА и Аа, имея одинаковый фенотип благодаря доминантности аллеля А над рецессивным аллелем а, обладают равной селективной ценностью перед лицом естественного отбора. Таким образом, под покровом доминантного аллеля А рецессивный аллель а может очень долго существовать в популяции даже в том случае, если гомозиготы по рецессивному аллелю аа имеют пониженную жизнеспособность (табл. 3).
Таблица 3. Темпы возрастания частоты доминантного аллеля А в популяции, испытывающей слабое действие отбора (особи с генотипами АА и Аа имеют 1%-ное селективное преимущество по сравнению с генотипом аа). По К. Пэтау, 1939
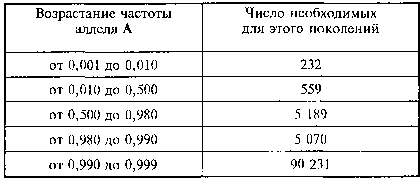
Как видно из теоретических расчетов, полученных в 1939 г. немецким теоретиком К. Пэтау[382], нарастание частоты доминантного аллеля в случае его селективной ценности вначале идет относительно быстро, но затем замедляется и не приводит на протяжении многих тысяч поколений к полному исчезновению селективно не выгодного рецессивного аллеля. При смене условий, в случае, если генотип аа будет иметь повышенную селективную ценность в сравнении с генотипами АА и Аа, повышение частоты аллеля а пойдет в обратном направлении в сравнении с рассмотренным в таблице и более быстрыми темпами. Это явление существования в популяции рецессивных аллелей в гетерозиготном состоянии было названо И. И. Шмальгаузеном и С. М. Гершензоном[383] в терминологии межвоенной поры «скрытым мобилизационным резервом внутривидовой изменчивости».
Генетикам-популяционистам того времени все более убедительным представлялось сведение микроэволюционного процесса к процессу постепенной смены генных частот, точнее, частот аллелей. Но поскольку и макроэволюция представлялась в ту пору бесспорным продолжением и развитием микроэволюции, то и вся эволюция стала постепенно восприниматься как процесс неуклонного и постепенного изменения частот аллелей. В результате к моменту возникновения синтетической теории эволюции генетикам-популяционистам стал представляться бесспорным процесс постепенности смены частот, т. е. постепенность стала казаться неотделимой чертой эволюционного процесса. Таким образом, произошел на новом уровне возврат к лейбницевскому «закону непрерывности», к дарвиновскому градуализму. Интересно, что на этот раз возврат к градуализму произошел в среде генетиков, среди которых еще за 20—30 лет до этого господствовали макромутационистские представления.
Однако градуализм преобладал лишь среди генетиков-популяционистов, притом связанных с изучением животных объектов. В 20—30-е годы стало ясно, что в видообразовании растений особую роль играет кратное увеличение числа хромосом — полиплоидия. В объединении идей эволюционной цитогенетики и ботаники участвовали исследователи многих стран. В СССР это были, в первую очередь, ученые школы Николая Ивановича Вавилова (рис. 204): Г. А. Левитский (рис. 205), от которого идут термины «кариотип» и «идеограмма»[384]; Г. Д. Карпеченко (рис. 206), впервые получивший плодовитый межродовой гибрид — рафанобрассику[385], плодовитость которого обеспечивалась за счет удвоения хромосомных наборов каждого из родительских видов; П. М. Жуковский и А. Р. Жебрак (рис. 207, 208), много сделавшие для изучения полиплоидии у злаков; М. А. Розанова, исследовавшая роль полиплоидии в природе у ягодных форм[386]; О. С. Стрелкова и А. П. Соколовская, на протяжении десятилетий изучавшие полиплоидию среди цветковых растений в природе и показавшие высокую концентрацию полиплоидов в экстремальных условиях высоких широт и высокогорий[387]; В. А. Рыбин, осуществивший ресинтез культурной сливы[388]; Дончо Костов — болгарский генетик, исследовавший полиплоидию у пасленовых, каучуконосов и других растений[389] и ресинтезировавший одну из форм табака; В. П. Зосимович, начавший исследование сахарной свеклы с изучения дикорастущих видов рода и полиплоидии у них[390]. Не меньший вклад внесли ученые школы Михаила Сергеевича Навашина[391] (Е. Н. Герасимова-Навашина, В. А. Поддубная-Арнольди, Н. А. Чуксанова), а также Лидия Петровна Бреславец[392]; в Швеции — А. Мюнтцинг[393], А. Густафсон[394], А. Леван[395], А. и Д. Лёвэ[396], в Японии — X. Кихара и его ученики[397]; в Дании — О. Хагеруп[398], в Германии — Э. Баур[399], Г. Тишлер[400] и Ф. Веттштайн[401]; в Англии — Сирил Дарлингтон (рис. 209)[402], создавший учение о хромосомах как генетических системах; большой коллектив американских цитогенетиков и ботаников — Э. Андерсон, Э. Бэбкок, Дж. Болдуин, А. Блексли, А. Эвери, Т. Гудспид[403], трио исследователей дикой флоры — Дж. Клаузен, Д. Кек и У. Хизи[404], а начиная с 40-х гг. ныне здравствующий выдающийся ботаник-эволюционист Л. Стеббинс (рис. 210)[405] — все они, и десятки других, не упомянутых здесь исследователей на конкретных примерах демонстрировали, в отличие от дрозофилистов, дискретность не только полиплоидии как цитогенетического механизма, но и видообразования в целом. Однако синтетическая теория эволюции (СТЭ) в большей степени создавалась исследователями животных, нежели растений, и этот огромный массив фактов из цитогенетики растений не был в полной мере аккумулирован СТЭ.

Рис. 204. Николай Иванович Вавилов.
Из: С. Резник (1968).

Рис. 205. Георгий Андреевич Левитский. Классик цитогенетики. Член-корреспондент АН СССР. Арестован в 1940 г. вместе с Н. И. Вавиловым. Погиб в лагерях.
Из: С. М. Гершензон (1992).

Рис. 206. Георгий Дмитриевич Карпеченко. Классик цитогенетики. Арестован в 1940 г. вместе с Н. И, Вавиловым. Погиб.
Из: С. М. Гершензон.

Рис. 208. Антон Романович Жебрак. Президент АН Белоруссии. В 1948 г. был снят со всех постов за критику Т. Д. Лысенко. В середине 50-х годов — заведующий кафедрой ботаники Московского фармацевтического института, где вместе с В. В. Сахаровым восстановил подготовку генетиков.
Из. Выдающиеся советские генетики (1980).

Рис. 207. Петр Михайлович Жуковский. Ботаник, генетик, эволюционист. Академик ВАСХНИЛ. Был директором вавиловского Всесоюзного института растениеводства. с 1948 г. был вынужден прекратить работы по генетике.
Из: Соратники Николая Ивановича Вавилова (1994).

Рис. 209. Сирилл Дарлингтон (1903-1981) — в центре, слева — Г. Г. Мёллер, справа — Н. В. Тимофеев-Ресовский. 1930-е гг.
Архив журнала «Природа».
Другим источником синтеза генетики и эволюционизма стали работы Н. И. Вавилова и его последователей по изучению закономерностей изменчивости. Вавиловский «закон гомологических рядов наследственной изменчивости» (1922) впервые в биологии открыл пути для прогнозирования фиксируемых естественным отбором мутаций. Там, где большинство исследователей видело простую случайность, Вавилов увидел закономерность. Эти работы Вавилова развивались не только его школой, но и вызвали интерес у таких далеких от изучения культурных растений биологов, как ихтиолог Л. С. Берг, как гистолог А. А. Заварзин-старший.

Рис. 210. Ледьярд Стеббинс.
Из: G. L. Stebbins (1952).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК