3. Заключительные замечания
Грибы S. pombe и N. crassa оказались мощными системами для обнаружения и объяснения эпигенетических явлений. Эпигенетика как область науки все еще находится в стадии младенчества, и эпигенетические механизмы продолжают появляться на свет. Поэтому не удивительно, что глубина и широта нашего сегодняшнего понимания таких эпигенетических процессов, как те, что описаны в настоящей главе, варьируют от организма к организму. Еще слишком рано судить о том, насколько общими являются описанные эпигенетические механизмы. Например, возможно, что для сайленсинга и формирования гетерохроматина некоторые организмы, подобно S. pombe, базируются главным образом на RNAi, тогда как другие, такие как N. crassa, основываются в большей степени на метилировании ДНК. Уже ясно, что даже эти два этих модельных эукариотных организма обладают и важными различиями, и существенным сходством. Оба организма используют метилирование гистона H3K9 и RNAi, при том что ни один из этих механизмов не обнаружен у почкующихся дрожжей, S. cerevisiae. Из этих трех грибов, однако, только Neurospora практикует метилирование ДНК. Стоит также заметить, что данный процесс может быть функционально довольно различным у этих двух организмов. Например, у Neurospora участие компонентов RNAi предполагается в «подавлении» и мейотическом сайленсинге, но не в формировании гетерохроматина, тогда как у дробянковых дрожжей компоненты RNAi вносят вклад в формирование гетерохроматина, но другие их роли не установлены. Наконец, следует отметить, что даже такие обшие черты, как гетерохроматин, ассоциированный с центромерами, у дробянковых дрожжей и у Neurospora могут обладать важными различиями. Важная цель на будущее состоит в том, чтобы открыть, в какой степени информация, собранная на одном организме, приложима к другим объектам. Дальнейшие исследования эпигенетических процессов у различных модельных организмов, в том числе S. pombe и N. crassa, дадут эту информацию. Мы надеемся, что грибы как чрезвычайно разнообразные организмы будут продолжать служить полезными системами для эпигенетических исследований на многие годы.
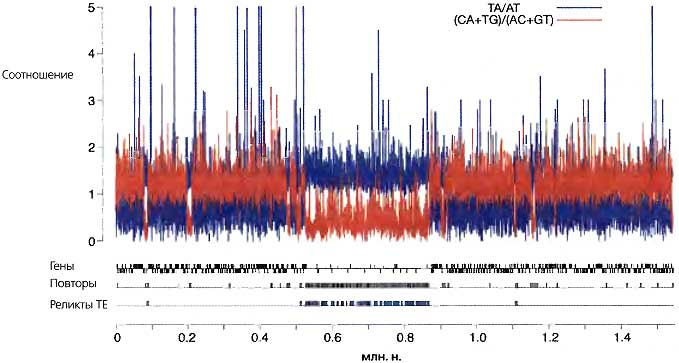
Рис. 6.16. Организация района центромеры 7 у N. crassa
Были собраны контиги 249,255 и 21 релиза 7.0 геномной последовательности (http://www.broad.mit.edu/annorntion/fungi-neurospora_crassa_7/index.html) и все кроме первых 400 т.о. последовательности контига 10, и этот объединенный файл последовательностей был проанализирован с инкрементом 200 п.н. на «индексы RIP» (ТрА/АрТ [синий цвет] и CpA+TpG/ApC+GpT [красный цвет]) (Galagan et al., 2003; Selker et al., 2003; Galagan and Selker, 2004). Район длиной -360 т.о. с высокой плотностью перемещаемых элементов (ТЕ), инактивированных RIP (остатки ретротранспозонов — синий цвет, остаток ДНК-транспозона — фиолетовый цвет), был обнаружен между маркерами, фланкирующими центромеру, которая была картирована генетически. Изображенный сегмент длиной -1.5 млн о. включает 383 аннотированных гена (над линией и под ней, чтобы показать гены в противоположных ориентациях), из которых только 20 коротких предсказанных гена находятся в пределах предсказанного центромерного района. Размеры перерывов в последовательности [sequence gaps] между контигами (позиции 0.5466, 0.6956 и 0.9058 млн о. на рисунке) неизвестны
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК