7. Фосфорилирование Н2АХ функционирует в репарации двунитевых разрывов ДНК
Гистоны Н2А также образуют семейство разных вариантов, обнаруживаемых у всех эукариот. Вариант Н2АХ определяется по присутствию мотива карбокситерминальной аминокислотной последовательности, SQ(E или D)?, где ? означает гидрофобную аминокислоту. Серии в этой последовательности-мотиве является сайтом фосфорилирования, продуцируя модифицированный белок, обозначаемый «?-Н2АХ». Динамическая природа хроматина и фосфорилирование Н2АХ особенно очевидны, когда в ДНК возникают двунитевые (ds, double-stranded) разрывы (Morrison and Shen, 2005). Летальность даже одиночного двунитевого разрыва требует немедленных действий по репарации повреждения и восстановления непрерывности двойной спирали. Обнаружение ds-разрыва в норме происходит в пределах минуты или около того с момента его образования, и это, в свою очередь, включает быстрое фосфорилирование Н2АХ в непосредственной близости от сайта разрыва. Это фосфорилирование выполняется членами семейства фосфоинозитол 3-киназоподобных киназ. Вслед за этим первоначальным событием фосфорилирование Н2АХ быстро распространяется вдоль по хромосоме, маркируя относительно большой хроматиновый домен, окружающий этот разрыв. Наконец, этот ds-разрыв в конце концов репарируется либо путем гомологичной рекомбинации, либо негомологичным воссоединением концов [nonhomologous end-joining], и метка-фосфорилирование удаляется.
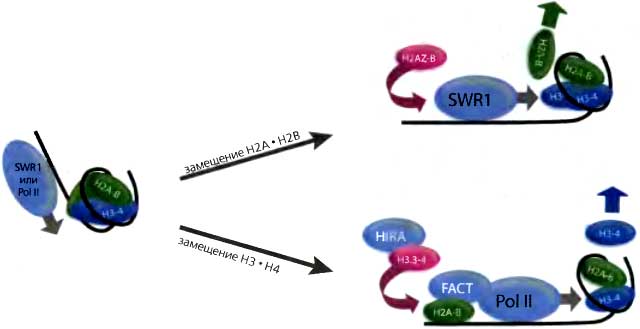
Рис. 13.7. Модель независимого от репликации замещения или обмена
Большая молекулярная машина (либо комплекс SWR1, либо РНК-полимераза II) частично или полностью расвертывает нуклеосому во время своего прохождения. Результатом этого оказывается либо сохранение гетеродимерных субъединиц, как, например, облегчаемый FACT перенос Н2А•Н2В из положения перед РНК-полимеразой в положение позади нее (Formosa et al. 2002; Belotserkovskaya et al. 2003), либо утрата гетеродимера. В последнем случае процесс репарации хроматина заменяет утраченный гетеродимер либо H2AZ•Н2В (верхняя часть рисунка), либо Н3.3•Н4 (нижняя часть рисунка)
Фосфорилирование Н2АХ не является существенным для обнаружения или репарации ds-разрывов, потому что делеция гена или мутация «мишени» — серинового остатка не отменяет репарацию. Однако Н2АХ не просто маркер повреждения, поскольку такие мутанты уменьшили эффективность репарации и являются гиперчувствительными к радиационному повреждению и генотоксическим агентам. В настоящее время полагают, что Н2АХ функционирует в процессе репарации ds-разрывов по крайней мере двумя способами. Во-первых, он может помогать рекрутировать или сохранять белки, требующиеся для репарации в сайте разрыва (Morrison and Shen, 2005). Во-вторых, он может стабилизировать хромосому, окружая разорванные концы, путем рекрутирования когезина, белкового комплекса, ответственного за удерживание вместе сестринских хроматид (Lowndes and Toh, 2005).
Эволюция Н2АХ не похожа на эволюцию других вариантов гистонов. Хотя ген Н2АХ обнаружен почти у всех эукариот, он возникал неоднократно в относительно недавнее время (Malik and Henikoff, 2003). Например, версия Н2АХ, обнаруженная у Drosophila, отличается от версии, обнаруживаемой у другого двукрылого насекомого, Anopheles Предположительно, способность развивать новый Н2АХ из канонической формы Н2А является следствием простого мотива SQ(E или D)©. Ожидается, что развитие такого мотива на карбоксильном конце канонического Н2А происходило неоднократно в ходе эволюции. Случайная утрата существующего Н2АХ с вновь отчеканенной версией могла бы подпитываться необходимостью для Н2АХ быть очень однородно распределенным, потому что ds-разрывы могут происходить в любом месте генома. Если в существующем гене Н2АХ происходят мутации, уменьшающие его сходство с каноническим Н2А таким образом, что его сборка становится менее эффективной или однородной, возникнет сильное давление отбора, чтобы заместить его версией, которая более похожа на канонический Н2А. Эта логика могла бы помочь объяснить исключительный случай Н2Ах Drosophila, который, в отличие от других эукариот, происходит не от своего канонического Н2А, а, скорее, от удаленной линии H2AZ (описываемой ниже). Если все, что нужно, чтобы быть Н2АХ — это находиться в положении Н2А в нуклеосоме и иметь карбокситерминальный мотив для фосфорилирования, H2AZ может развить в себе эти способности.
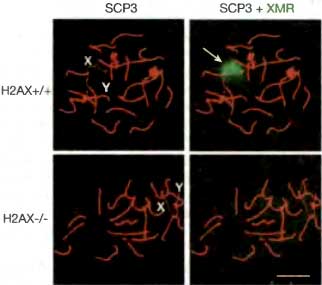
Рис. 13.8. Стадия пахитены сперматогенеза, показывающая зависимость формирования полового тельца от Н2АХ
В нормальных сперматоцитах млекопитающих видно, что ядерная структура, называемая половым тельцем (стрелка, помечено зеленым цветом на правых рисунках), охватывает неспаренный бивалент XY (помечен на левых рисунках). Синаптонемный комплекс, выравнивающий спаренные хромосомы, окрашен в красный цвет. Половое тельце в норме обогащено Н2АХ (Н2АХ+/+). В сперматоцитах Н2АХ-/- половое тельце не образуется, и эпитоп полового тельца становится диспергированным по аутосомам (нижний правый рисунок). Масштабная линейка = 10 мкм. Фотографии любезно предоставлены Shantha Mahadevaiah и Paul Burgoyne (Fernandez-Capetillo et al., 2003)
Репарация ds-разрывов представляет собой несомненно универсальную функцию фосфорилирования Н2АХ, и, казалось бы, у этого процесса нет никакого стабильного эпигенетического аспекта. Однако мыши с нуль-мутацией по Н2АХ стерильны, и цитологическое изучение сперматогенеза млекопитающих выявило яркую эпигенетическую особенность — специфическое фосфорилирование Н2АХ на биваленте XY (рис. 13.8) (Femandez-Capetillo et al., 2003). Эта хромосомная пара занимает отдельное «половое тельце» во время мейотической профазы, предположительно участвующее в сайленсинге сцепленных с полом генов во время мужского мейоза. Фосфорилирование Н2АХ играет существенную роль в формировании нормального полового тельца, и дефектные по Н2АХ сперматоциты не способны спариваться или конденсироваться и не могут инактивировать гены X и Y во время мейоза. Фосфорилирование Н2АХ бивалента XY отличается от процесса, происходящего в ds-разрывах. Фосфорилирование XY в половом тельце не требует разрывов; скорее, оно происходит наиболее заметным образом в неспаренных участках хромосом. Механизмы, посредством которых фосфорилирование Н2АХ «нацеливается» на неспаренные хромосомы, и то, как это событие приводит к конденсации, спариванию и сайленсингу, в настоящее время неизвестны. Однако интересно пофантазировать, что эта роль может иметь отношение к способности взаимодействовать с когезином и рекрутировать его.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК