4.3. Способность стволовых клеток к репрограммированию
Постоянное поддержание плюрипотентности в культуре без признаков старения вполне возможно требует постоянного эпигенетического репрограммирования стволовых клеток. То, что эти клетки имеют репрограммирующую активность, было показано в экспериментах по слиянию EG или ES-клеток с дифференцированными соматическими клетками (Tada et al., 1997, 2001; Cowan et al., 2005). Было установлено, что в тетраплоидных клеточных линиях, получившихся в результате слияния, соматический эпигенотип был репрограммирован (рис. 20.9). При слиянии EG и соматических клеток соматический геном утрачивал ДНК метилирование импринтных генов, а также других последовательностей в геноме (Tada et al., 1997). Напротив, при слиянии ES-клеток с соматическими ДНК метилирование импринтных генов сохранялось, но инактивированная X хромосома (в женских клетках) реактивировалась, промотор гена Oct4 становился ДНК деметилированным, что приводило к реэкспрессии Oct4 (Tada et al., 2001; Cowan et al., 2005; Surani, 2005).
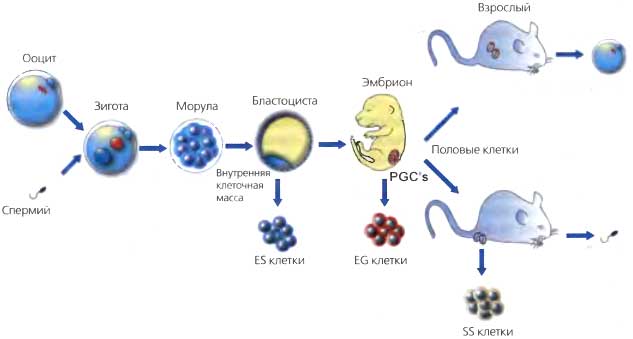
Рис. 20.8. Связь между развитием половых клеток и плюрипотентностью
Ооцит и зигота, а также клетки морулы до определенной стадии являются тотипотентными (ооцит имеет ограничения, налагаемые родительским импринтингом). Плюрипотентные клетки можно получить из клеток внутренней клеточной массы (ICM), примордиальных половых клеток (PGC) и сперматогониальных стволовых клеток (SS) новорожденных и взрослых семенников. Таким образом, существует тесная связь между половыми клетками в процессе развития и плюрипотентностью
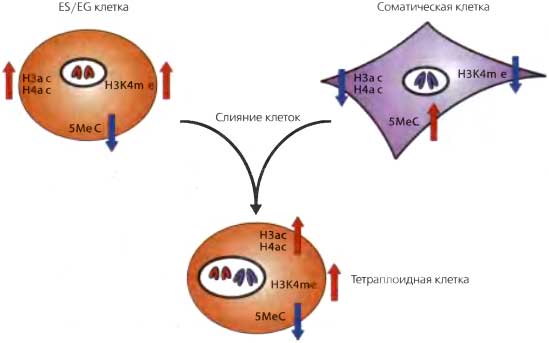
Рис. 20.9. Плюрипотентные клетки обладают способностью репрограммировать соматические клетки ES- или EG-клетки могут сливаться с соматическими клетками с образованием тетраплоидных клеток. Это приводит к эпигенетическому репрограммированию соматических ядер, в которых происходят изменения, например, в 5шеС, H3ас, Н4ас и H3K4me. Тетраплоидные ядра в результате слияния и репрограммирования приобретают плюрипотентный фенотип: при инъекции в бластоцисты они вносят вклад в различные клеточные типы эмбриона
Факторы репрограммирования, влияющие на ДНК метилирование и модификацию гистонов в этих гибридах, пока неизвестны. Например, неясно, требует ли деметилирование в EG или ES клетках активности ДНК деметилазы или репликация ДНК происходит в отсутствие Dnmtl, в результате чего происходит пассивное деметилирование. Тем не менее, ES- и EG-клетки должны рассматриваться как важный источник для выделения и характеристики эпигенетических факторов репрограммирования.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК