3.2. Регуляция импринтированной Х-инактивации
Отцовски импринтированную Х-инактивацию впервые наблюдали у сумчатого (Sharman, 1971). Впоследствии Такаги и Сасаки (Takagi and Sasaki, 1975) продемонстрировали импринтированную Х-инактивацию в экстра-эмбриональных клеточных линиях ТЕ и РЕ мышиных эмбрионов. Именно происхождение Х-хромосомы от того или другого родителя управляет ее статусом; другими словами, инактивируются отцовские, но не материнские Х-хромосомы, независимо оттого, сколько Х-хромосом или хромосомных наборов присутствуют в клетке. Обратите внимание на то, что единственная Х-хромосома у самцов XYвсегда происходит от матери и, следовательно, не инактивируется в импринтированных тканях.
Какова же тогда природа импринта? Исследования экспрессии Xist, регулирующего Х-инактивацию в cis-конфигурации, показывают, что у мышиных эмбрионов имеется репрессивный импринт на аллели, локализованной в Xm (рис. 17.4). Этот импринт препятствует экспрессии Xist, удерживая Х-хромосому в активном состоянии. Опыты с пересадкой ядер показали, что этот репрессивный А75/-импринт устанавливается во время созревания ооцитов (Tada et al., 2000). Молекулярная основа этого импринта неизвестна, но метилирования ДНК не требуется, в отличие от многих других импринтированных генов (см. детали геномного импринтинга в главе 19).
Одна из теорий предпочтительной инактивации Хр в зиготе заключается в том, что имеет место перенос сайленсинга бивалента XY, который устанавливается на стадии пахитены мейоза у самцов (meiotic sex chromosome /motivation, MSCI) (Huynh and Lee, 2003). Недавние исследования говорят против этого. Во-первых, было показано, что MSCI — отдельный и не зависимый от Xist механизм, включаемый в пахитене присутствием неспаренных участков хромосом, как на половых хромосомах, так и на аутосомах (Turner et al., 2005 и помещенные там ссылки). Во-вторых, анализ экспрессии ряда сцепленных с X генов в ранних зиготах показал, что сайленсинг Хр возникает de novo в ответ на экспрессию Xist зиготической Хр (Okamoto et al., 2005 и помещенные там ссылки).
Экспрессия отцовского Xist (и получающийся в результате этого сайленсинг Хр), начинающаяся в начале зиготической активации гена (на 2—4-клеточной стадии), показывает, что аллель Xist хромосомы Хр готова к экспрессии (рис. 17.4). Однако в мужских соматических тканях Xist всегда репрессирована, откуда следует, что мужские зародышевые клетки должны каким-то образом ремоделировать локус Xist. В соответствии с этим, имеет место специфичное в отношении района деметилирование сайтов CpG в промоторе Xist во время сперматогенеза (Norris et al., 1994).
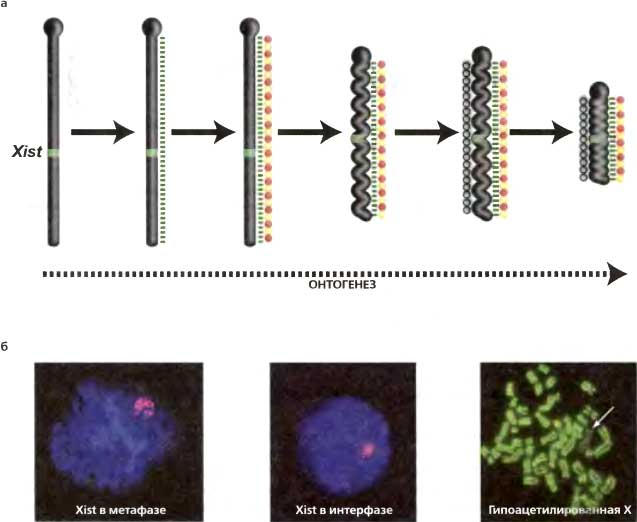
Рис. 17.3. Прогрессивная гетерохроматинизация всей хромосомы, индуцируемая Xist-РНК
(а) Когда ген Xist экспрессируется, синтезируемая РНК связывается с Х-хромосомой, с которой она транскрибируется, и покрывает ее (зеленая прерывистая линия). Считается, что Xist-РНК включает сайленсинг этой хромосомы, рекрутируя активности, модифицирующие хроматин (красные и желтые кружки). Эта первоначальная волна сайленсинга, в свою очередь, приводит к рекрутированию дополнительных слоев эпигенетической модификации (серые кружки), еще более стабилизируя структуру гетерохроматина. Установление этих различных уровней эпигенетического сайленсинга достигается поэтапно в ходе всего развития и онтогенеза. (б) Локализацию Xist-PHК по всем Х-хромосомам показывает гибридизация in situ как в интерфазе, так и в метафазе
У эмбрионов с анеуплоидией по Х-хромосоме результатом родительских импринтов, управляющих экспрессией Xist, оказываются неправильные паттерны Х-инактивации на ранних этапах развития. Так же обстоит дело у андрогенетических и гиногенетических (или партеногенетических) эмбрионов, у которых оба хромосомных набора происходят, соответственно, либо от отца, либо от матери. Хотя это обычно снижает жизнеспособность эмбриона, некоторые эмбрионы выживают, очевидно исправляя несоответствующие паттерны Х-инактивации, чтобы обеспечить наличие одной активной Х-хромосомы во всех клетках. Полагают, что это связано с преодолением импринта с помощью механизма, в норме регулирующего случайную Х-инактивацию (см. ниже).
Ген Tsix, антисмысловой регулятор Xist, необходим для импринтированной Х-инактивации, поскольку делеция главного промотора приводит к ранней эмбриональной летальности, когда передается материнской, но не отцовской гаметой (Lee, 2000). Летальность, по-видимому, можно приписать несоответствующей экспрессии Xist хромосомы Xm, т. е. неспособности сохранить активную Х-хромосому у эмбрионов как XmY, так и ХгпХр. В настоящее время неясно, является ли экспрессия Tsix-РНК первичным импринтом или же функционирует позже для поддержания импринта.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК