8. Комплексы, осуществляющие ремоделинг хроматина, и варианты гистонов
Другим важным механизмом, посредством которого индуцируются переходы в хроматиновой матрице, является выработка сигналов [signaling] для рекрутирования комплексов «ремоделинга» хроматина, использующих энергию (гидролиз АТФ) для изменения хроматина и состава нуклеосом нековалентным образом. Нуклеосомы, особенно когда они связаны репрессивными факторами, ассоциированными с хроматином, нередко «навязывают» транскрипционной машине состояние значительного подавления. Отсюда лишь некоторые транскрипционные факторы и регуляторы, специфичные к определенным последовательностям (хотя и не базовая транскрипционная машина), способны получать доступ к сайту (сайтам) своего связывания. Эта проблема доступа решается, хотя бы отчасти, белковыми комплексами, которые мобилизуют нуклеосомы и (или) изменяют нуклеосомную структуру. Ремоделинг хроматина часто функционирует совместно с ферментами, активирующими модификации хроматина, и связанные с ним активности в целом можно разделить на два семейства: семейство SNF2H, или ISWI, и семейство Brahma, или SWI/SNF. Семейство SNF2H/ ISWI мобилизует нуклеосомы вдоль ДНК (Tsukiyama et al., 1995: Varga-Weisz et al., 1997), тогда как Brahma/SWI/ SNF на время изменяет структуру нуклеосомы, экспонируя контакты ДНК: гистон с использованием механизмов, которые только сейчас начинают раскрываться (глава 12).
Кроме того, некоторые из гидролизующих АТФ активностей схожи с «обменными комплексами», которые сами предназначены для замещения обычных коровых гистонов специализированными «вариантными» гистоновыми белками. Эта осуществляемая с затратами АТФ перетасовка в действительности может быть средством замещения существующих модифицированных гистоновых «хвостов» новым, свободным от старого, набором вариантных гистонов (Schwartz and Ahmad, 2005). Альтернативная возможность заключается в том, что рекрутирование таких комплексов ремоделинга хроматина, как SAGA (Spt-Ada-Gcn5-aцeтилтpaнcфepaзa), может быть также усилено пред существующими модификациями гистонов, чтобы обеспечить транскрипционную компетентность промоторов-мишеней (Grant et al., 1997; Hassan et al., 2002).
В дополнение к инициации транскрипции и установлению первичного контакта с промоторным районом прохождению РНК-полимеразы II (или РНК-полимеразы 1) во время происходящей в ходе транскрипции элонгации препятствует, сверх того, присутствие нуклеосом Поэтому требуются механизмы для обеспечения завершения образующихся транскриптов (особенно с длинных генов). В частности, ряд гистоновых модификаций и докинг-эффекторов действуют совместно с такими комплексами ремоделинга хроматина, как SAGA и FACT (для облегчения транскрипции хроматина) (Orphanides et al., 1998), обеспечивая прохождение РНК-полимеразы II через нуклеосомные порядки. Эта совместная активность обычно индуцирует, например, увеличенную мобильность нуклеосом, смещает димеры Н2А/Н2В и стимулирует обмен коровых гистонов на гистоновые варианты. Как таковая, она дает прекрасный пример тесного взаимодействия между модификациями гистонов, ремоделингом хроматина и обменом на гистоновые варианты для облегчения инициации транскрипции и последующей элонгации (Sims et al., 2004). Были охарактеризованы и другие комплексы ремоделинга, такие как Mi-2 (Zhang et al., 1998; Wade et al., 1999) и INO-80 (Shen et al., 2000), участвующие в стабилизации репрессированного, а не активного хроматина.
Композиционные различия хроматиновой фибриллы, которые возникают благодаря присутствию гистоновых вариантов, вносят вклад в индексирование участков хромосом для специальных функций Каждый гистоновый вариант представляет собой замену для конкретного корового гистона (рис. 3.12), хотя гистоновые варианты часто составляют малую долю от всей массы гистонов, в силу чего их труднее исследовать, чем регулярные гистоны. Возрастающий объем литературы (обзор см.: Henikoff and Ahmad, 2005; Sarma and Reinberg, 2005) свидетельствует, что гистоновые варианты обладают своей собственной характеристикой восприимчивости к модификациям, вероятно определяемой небольшим числом аминокислотных замен, отличающих их от членов их семейства. С другой стороны, некоторые варианты гистонов имеют отличающиеся амино- и карбокситерминальные домены с уникальной, в отношении регуляции хроматина, активностью и различным сродством к связывающимся факторам. В качестве примера можно привести транскрипционно активные гены, у которых гистон H3 заменен вариантом H3.3, в связанном с транскрипцией механизме, который не требует репликации ДНК (Ahmad and Henikoff, 2002). Замещение корового гистона Н2А вариантом H2A.Z коррелирует с транскрипционной активностью и может индексировать 5 — конец свободных от нуклеосом промоторов. Однако H2A.Z был ассоциирован также и с репрессированным хроматином. CENP-A, специфичный для центромеры вариант гистона H3, существен для центромерной функции и, отсюда, для сегрегации хромосом. Н2А.Х, вместе с другими гистоновыми метками, ассоциируется с выявлением повреждений ДНК и, по-видимому, индексирует повреждение в ДНК для рекрутирования комплексов репарации повреждений ДНК. МакроН2А — вариант гистона, специфически ассоциирующийся с неактивной Х-хромосомой (Xi) у млекопитающих (дальнейшие детали по вариантам гистонов см. в главе 13).
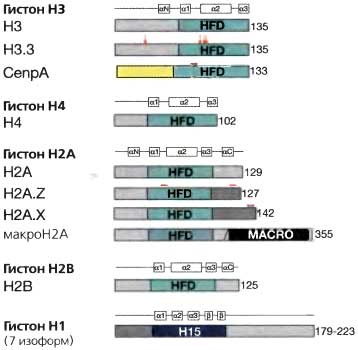
Рис. 3.12. Варианты гистонов
Доменная структура белка для коровых гистонов (H3, Н4, Н2А, Н2В), линкерного гистона Н1 и вариантов гистонов H3 и Н2А. Показаны свернутый домен гистона (HFD — histone fold domain), где происходит димеризация гистона, и участки белка, отличающиеся у гистоновых вариантов (показаны красным цветом)
Важно отметить (и это противоречит обычно встречающемуся в учебниках утверждению, что гистоны синтезируются и занимают свое место только во время S-фазы), что синтез и замещение многих из этих гистоновых вариантов происходят независимо от репликации ДНК. Следовательно, замещение коровых гистонов гистоновыми вариантами не ограничено стадиями клеточного цикла (т. е. S-фазой), но может немедленно вступать в силу в ответ на действующие в данный момент механизмы (например, транскрипционную активность или напряжение кинетохора во время клеточного деления) или стрессовые сигналы (например, повреждение ДНК или голодание). Элегантные биохимические исследования позволили документировать ремоделинг хроматина или обменные комплексы, специфичные для замещения отдельных гистоновых вариантов, таких как H3.3, H2A.Z или Н2А.Х (Cairns, 2005; Henikoff and Ahmad, 2005; Sarnia and Reinberg, 2005). Например, замещение H3 вариантом H3.3 опосредуется действием обменного комплекса HIRA (регулятор гистона A) (Tagani et al., 2004), а Н2А замещается H2A.Z благодаря активности обменного комплекса SWR1 (связанная с Swi2/ Snf2 АТФаза I) (Mizuguchi et al., 2004). В совокупности эти замещения позволяют вариантным нуклеосомам строить особенно активный хроматин. Для некоторых других гистоновых вариантов механизм «нацеливания» и обмена еще остается определить; они осуществляются либо через зависимый от АТФ комплекс обмена гистонов, либо через шаперонный белок гистонов Сейчас даже постулировали, что обменные комплексы могут существовать для замещения модифицированных гистонов их модифицированными партнерами как механизм для стирания более прочных эпигенетических меток, размещенных в аминоконцах гистонов.
Метилирование ДНК — эпигенетический механизм, о котором раньше других стало известно, что он коррелирует с репрессией генов (Razin and Riggs, 1980). В той
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК