1.4. Центромерные внешние повторы без посторонней помощи делают возможной сборку «молчащего» хроматина
Зависимый от Clr4 «молчащий» хроматин собирается не только в центромерах, но и над районом размером около 20 т.о., содержащим «молчащие» локусы типа спаривания (mat2-mat3) (Noma et al., 2001) и соседствующим с терминальными теломерными повторами (консенсус TTACAGG), добавляемыми теломеразой (Nimmo et al., 1994, 1998; Allshire et al., 1995; Kanohetal., 2005). Район cenH (т.е. гомологичный центромере), который находится "Между mat2 и mat3, имеет большое сходство, свыше ~7 т.о., с внешними повторами, обнаруживаемыми в центромерах (Grewal and Klar, 1997). Кроме того, по крайней мере 0.5 т.о. нуклеотидных последовательностей ДНК, имеющих >84 % идентичности с повторами cenH, могли бы действовать в cis-конфигурации, осуществляя сборку «молчащего» хроматина.
Сборка зависимого от Clr4 «молчащего» хроматина происходит даже на соседних маркерных генах, когда центромерный внешний повтор (dg) или нуклеотидные последовательности mat2-mat3 (cenH) вставляются в районы генома, где в норме сайленсинг не происходит («эктопический» сайленсинг; рис. 5) (Ayoub et al., 2000; Partridge et al., 2002; Volpe et al., 2003). Простое объяснение могло бы заключаться в том, что связывающиеся с ДНК белки узнают эти повторы и, когда связываются, эти белки рекрутируют HDACs и HKMT Clr4, результатом чего является метилирование H3K9, связывание хромодоменных белков и формирование гетерохроматина. Однако ситуация на самом деле еще более сложная. Теперь известно, что центромерные внешние повторы транскрибируются. Замечательно, что эта транскрипция приводит к образованию двухнитчатой РНК (dsRNA) — субстрата для RNAi-машинерии, — а она затем рекрутирует Clr4 HKMT для включения сборки «молчащего» хроматина (Volpe et al., 2002; Sadaie et al., 2004). RNAi также действует на родственные повторы cenH, обнаруживаемые в районе mat2-mat3 и в повторах TAS в теломерах (Cam et al., 2005; Kanoh et al., 2005).
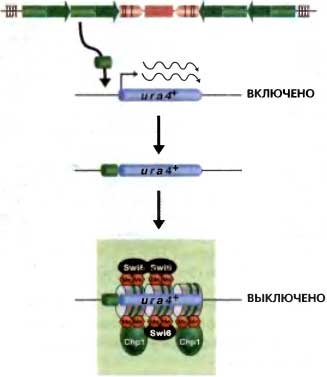
Рис. 6.5. Нуклеотидные последовательности центромерных повторов опосредуют сайленсинг
Вставка фрагментов (1—2 т.о.) рядом с экспрессируемым геном ura4+ в локусе, который в норме не склонен к сайленсингу, приводит к метилированию НЗК9, связыванию Swi6 и Chp1 и транскрипционному сайленсингу
Однако картина еще более усложняется тем обстоятельством, что в mat2-mat3 RNAi действует, устанавливая «молчащий» хроматин, в то время как связывающиеся с ДНК белки Arf1 и Perl связываются вблиз cenH и поддерживают «молчащий» хроматин над mat2-mat3 даже в отсутствие ключевых компонентов RNAi (Jia et al., 2004; Kim et al., 2004). Аналогично этому, перекрывающиеся механизмы сайленсинга действуют и в теломерах; терминальные повторы без посторонней помощи могут рекрутировать Clr4 HKMT и. таким образом, Swi6 через связывающийся с теломерными повторами белок Tazl, но RNAi действует также через cenH-часть элементов TAS, формируя расширенный район «молчащего» хроматина в теломерах (Nimmo et al., 1994; Allshire et al., 1995; Kanoh et al., 2005). Существует ли сравнимый перекрывающийся механизм для поддержания «молчащего» хроматина в центромерах? Хотя в клетках, лишенных RNAi, сайленсинг репортерных генов внешних повторов и самих транскриптов центромерных внешних повторов является дефектным, H3K9me2 сохраняется на хроматине внешних повторов в отсутствие компонентов RNAi (Sadaie et al., 2004). Какие факторы отвечают за поддержание этого метилирования? Одна возможность заключается в том, что белки, родственные CENP-B (Abpl, Cbhl, Cbh2), содержащие консервативный связывающийся с ДНК домен, связываются с ДНК центромерных повторов и являются необходимыми для эффективного формирования «молчащего» хроматина (Irelan et al., 2001; Nakagawa et al., 2002) Другие наблюдения показывают, что HDAC Clr4 также действует независимо от RNAi, поддерживая целостность гетерохроматина (Yamada et al., 2005).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК