4. Гетерохроматин и мейотическое спаривание / расхождение
Меотический процесс у большинства организмов охватывает (1) спаривание, которое выравнивает гомологи относительно друг друга; (2) синапсис, которое соединяет гомологи структурой, известной как синаптонемный комплекс (SC); (3) рекомбинацию, которая физически сцепляет гомологичные хромосомы и осуществляет обмен генетической информацией; (4) расхождение гомологов к противоположным полюсам в мейозе I и (5) разделение сестринских хроматид в мейозе II (рис. 14.9). Такие гетерохроматиновые элементы, как центромеры и теломеры, играют важную роль в контроле положения рекомбинационных событий в эухроматине; и наверняка гетерохроматиновые элементы (в особенности центромера) играют критическую роль в расхождении Более того, многочисленные недавние исследования убедительно свидетельствуют, что модификации гистонов и, возможно, другие эпигенетические компоненты и процессы играют критическую роль в облегчении мейотического процесса. Например, белок HIM-17 у С. elegans, который нужен для метилирования H3K9, необходим для формирования DSBs, инициирующих рекомбинацию (Reddy and Villeneuve, 2004). Аналогичным образом, для инициации DSBs у S. pombe требуется киназа гистонов (Ogino et al., 2006). Метилирование гистонов также необходимо, чтобы сделать мейотические хромосомы компетентными к завершению мейотических делений у Drosophila (Cullen et al., 2005; Ivanovska et al., 2005) и млекопитающих (De La Fuente, 2006).
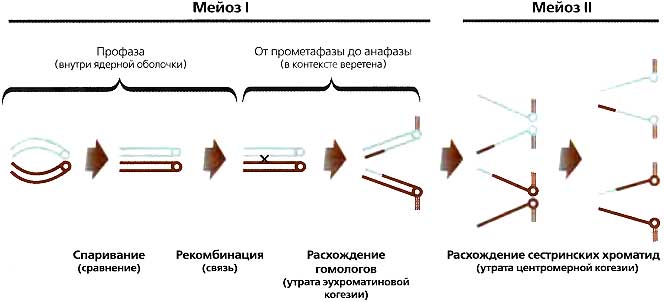
Рис. 14.9. Механистическое представление мейотического процесса
Пара гомологичных хромосом во время первого мейотического деления должна совершить три вещи. Во-первых, гомологи должны спариться по всей своей длине. Практически у всех организмов кульминацией этого спаривания является интимная ассоциация, при которой эти гомологи соединены вдоль всей своей длины синаптонемным комплексом. Это состояние называется синапсисом. Во-вторых, практически у всех организмов гомологичные хромосомы соединяются вместе рекомбинацией, называемой также кроссинговером. События обмена (кроссоверы) формируют структуры, называемые хиазмами, которые физически соединяют хромосомы друг с другом посредством сестринской хроматиды, представленной на каждом гомологе на обеих сторонах кроссоверного события. Как спаривание, так и рекомбинация происходят во время профазы (до разрушения ядерной оболочки). Третье важное событие, расхождение (сегрегация), происходит на MI-веретене, которое создается после разрушения ядерной оболочки. В ходе ранних стадий сборки веретена (прометафаза) хромосомы собираются вместе и образуют метафазную пластинку. У большинства животных самцы содержат центриолярные мейотические веретена, тогда как у самок большинства животных веретено ацентриолярно. В этом случае сами хромосомы образуют массу, на месте которой в конечном счете формируется метафазная пластинка, и организуют вокруг нее биполярное веретено. Коль скоро хромосомы должным образом совместно ориентированы (т.е. уравновешены в метафазной пластинке гомологичными кинетохорами, прикрепленными к противоположным полюсам веретена), ряд механизмов включают начало анафазы. В анафазе соединение (когезия) сестринских хроматид ослабляется вдоль плеч (но не возле центромер). Это растворяет связи, называемые хиазмами и созданные кроссоверами, и таким образом дает гомологам возможность разделиться и отойти к противоположным полюсам в анафазе I. Мейоз II в основе своей является гаплоидным митозом, который происходит и без репликации, и без рекомбинации
Хотя использование рекомбинации для обеспечения расхождения мейотических хромосом является почти универсальным, существуют также мейотические системы, в которых ассоциации гомологов могут устанавливаться без рекомбинации (т.е. так называемые ахиазматические мейотические системы). В таких системах гетерохроматиновые спаривания и, возможно, другие субстраты для эпигенетических модификаций оказываются играющими критическую роль. Это в особенности верно в отношении самок и самцов Drosophila melanogaster, но может быть верным и по отношению к дрожжам. Наконец, известны многие системы мейотического драйва — например, благоприятствование одной из аллелей в ходе мейотической сегрегации, — в которых модификация специфических гетерохроматиновых элементов вызывает или делает возможной последующую утрату инактивации специфической хромосомы или целого хромосомного набора.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК