1. Генетические и молекулярные методы исследования дрожжей
Дрожжи представляют собой гибкую и быструю генетическую систему для изучения клеточных процессов.
При времени клеточной генерации около 90 мин. всего за два дня можно вырастить колонии, содержащие миллионы клеток. К тому же дрожжи можно поддерживать в гаплоидной и диплоидной формах, что резко упрощает их генетический анализ. Как и бактерий, гаплоидные клетки дрожжей можно использовать для получения ауксотрофных мутантов со специфическими требованиями к составу питательной среды. Рецессивные летальные мутации можно поддерживать в культуре гаплоидных клеток в виде условных летальных аллелей (например, температуро-чувствительных мутантов) или в культуре гетерозиготных диплоидных клеток (содержащих и аллель дикого типа и мутацию). Наличие высокоэффективной системы гомологичной рекомбинации у дрожжей позволяет произвольно изменять любую выбранную хромосомную последовательность ДНК. К тому же, можно осуществлять манипуляции с участками хромосом в составе рекомбинантных плазмид, которые стабильно поддерживаются в делящихся клетках дрожжей, благодаря включению в них коротких последовательностей центромеры и ориджина репликации ДНК. В дрожжах стабильно поддерживаются даже линейные плазмиды (минихромосомы), содержащие теломерные повторы по концам.
Явление мозаичности, обусловленной эффектом положения, на модели репортерного гена white дрозофилы сыграло важную роль в изучении эпигенетической регуляции генов, а также генов, влияющих на эту уникальную форму репрессии (см. более детально в главе 5). Открытию и изучению аналогичного явления в области теломер у дрожжей, теломерного эффекта положения (telomere position effect — ТРЕ) помогло использование в качестве репортерных генов Ura3 и Ade2 (рис. 4.1). В присутствии 5-фтороротовой кислоты (5-fluoroorotic acid-5-FOA) продукт гена Ura3 превращает ее в 5-фторурацил (5-fluorouracil-5-FU) — ингибитор синтеза ДНК, вызывающий гибель клеток Однако, при интеграции в область гетерохроматина ген Ura3 оказывается репрессированным в части клеток, и именно эти клетки оказываются способными расти на среде с 5-FOA. Таким образом, определяя эффективность роста на среде с 5-FOA с помощью серийных разведений культуры клеток дрожжей (рис. 4.1а), можно количественно измерять степень репрессии репортерного гена в очень широких пределах (например, в 10— 106-кратных). Более того, мутации, нарушающие ТРЕ, могут быть легко идентифицированы по повышению чувствительности к 5-FOA.
Аналогично, при интеграции гена Ade2 в область гетерохроматина происходит его репрессия, и в клетке накопливается предшественник биосинтеза аденина, окрашивая ее в красный цвет Существенно, что эпигенетическая природа репрессии Ade2 видна в пределах одиночной колонии генетически идентичных клеток. Ген может быть «включен» в некоторых клетках и «выключен» в остальных. Внешне это проявляется как наличие красных участков на фоне белой колонии или наоборот (рис. 4.16). В отличие от теста с использованием Ura3, селекции против клеток, в которых остался не репрессированным ген Ade2, не происходит, поэтому фенотип клеток с интегрированным в субтеломерный гетерохроматин геном-репортером можно использовать как показатель скорости переключения и наследования эпигенетического состояния. Цветной тест с использованием Ade2 представляет собой удивительную иллюстрацию полустабильной природы репрессированного и дерепрессированного состояний.
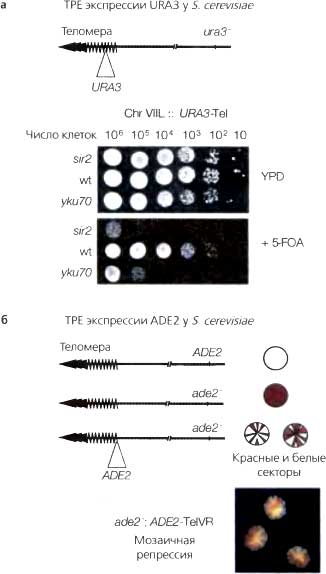
Рис. 4.1. Сайленсинги ТРЕ у дрожжей
(а) Ген Ura3, встроенный рядом с простыми TG-богатыми теломерными повторами в левом плече VII хромосомы, подвергается сайленсингу теломерным гетерохроматином в данном штамме дрожжей. При выращивании на обычной богатой среде (YPD) не наблюдается различий между клетками дикого типа (wt), клетками с репрессированным субтеломерным геном Ura3 и мутантами по сайленсингу, у которых теломерный гетерохроматин утрачен, а ген Ura3 экспрессируется. На среде, содержащей 5-FOA (нижняя панель), с другой стороны, клетки с репрессированным геном Ura3 (например, клетки дикого типа) могут расти, а клетки, экспрессирующие его (sir2nyku70 мутанты), не могут. Это объясняется тем, что продукт гена Ura3 превращает 5-FOA в токсичный интермедиат 5-фторурацил. Тест с серийными разведениями позволяет детектировать события сайленсинга даже при частоте 1 на 106 клеток.
(б) Клетки, содержащие ген Ade2 дикого типа, дают «белую» колонию, а клетки с мутантным ade2 — красную, что связано с накоплением красного интермедиата биосинтеза аденина. При встраивании гена Ade2 рядом с теломерой в правом плече V хромосомы происходит его эпигенетический сайленсинг. И «молчащее», и активное состояния гена Ade2 наследуются в генетически идентичных клетках, в результате чего образуются колонии с белыми и красными секторами (весьма похоже на картину, наблюдаемую при PEV)
Вместе с описанными генетическими подходами к протеазо-дефицитным штаммам, растущим в синхронных и асинхронных крупномасштабных культурах, вполне применимы биохимические методы. В последние годы арсенал доступных средств расширился изощренными методами анализа с помощью микрочипов и белковых сетей, легко охватывающих весь небольшой геном дрожжей. Эти методы позволяют осуществлять полногеномный анализ транскрипции, связывания факторов транскрипции, модификации гистонов и белок-белковых взаимодействий. Этот широкий арсенал изощренных методов позволил ученым исследовать механизмы регуляции формирования гетерохроматина и его физиологическую роль в клетках дрожжей. Однако, прежде чем описать эти открытия, необходимо более детально рассмотреть жизненный цикл дрожжей.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК