13. Старение и Sir2 связаны нестабильностью повторов РДНК
У дрозофилы активные повторы рДНК расположены по соседству с центромерным гетерохроматином, а у многих высших эукариот ядрышки и конденсированный гетерохроматин расположены рядом друг с другом. Следовательно генетическая и физическая ассоциация белка Sir2 с активно транскрибирующимися повторами рДНК, не зависящая от других SIR-белков, у дрожжей имеет существенное функциональное значение (Gotta et al., 1997). И действительно, Sir2 подавляет рекомбинацию рДНК (Gottlieb and Esposito, 1989) и репрессирует транскрибируемые PH К-полимеразой II гены-репортеры, встроенные в область рДНК-повторов. Из-за своей тандемно повторенной организации рДНК склонна к процессам неэквивалентной рекомбинации, приводящим к изменению числа ее повторов. С такой нестабильностью связаны и процессы возникновения кольцевых внехромосомных молекул рДНК (рис. 4.11) (Sinclair and Guarente, 1997). Sir2 требуется и для репрессии репортерных генов, встроенных в рДНК, и для предотвращения аберрантной рекомбинации, ведущей к потере повторов рДНК. Вероятными механизмами его действия являются позиционирование нуклеосом (Fritze et al., 1997) и выравнивание сестринских хроматид относительно друг друга, предотвращающее события неравного обмена (Kobayashi et al., 2004).
Наиболее неожиданным фенотипическим эффектом нуль-мутаций Sir2 является уменьшение продолжительности жизни, которое у дрожжей не связано напрямую с нарушением теломерной репрессии и уменьшением длины области TG-повторов. Короткоживущий фенотип дефектных по Sir2 дрожжей проявляется в том, что они проходят в среднем менее 12 клеточных делений по сравнению с 20—25 у дрожжей дикого типа (Kaeberlein et al., 1999). В настоящее время твердо установлено, что образование и накопление внехромосомных кольцевых молекул рДНК (ERC) в результате неэквивалентной рекомбинации у дрожжей коррелирует со старением (рис. 4.11). Важно отметить не только то, что при потере Sir2 уменьшается продолжительность жизни дрожжей, но и что при суперэкспрессии Sir2, сопровождающейся увеличением количества его комплексов с рДНК, продолжительность жизни увеличивается. Другие мутации, уменьшающие эффективность вырезания рДНК, например элиминация барьерного белка репликативных вилок Fobl (Defossez et al., 1999), также увеличивают продолжительность жизни дрожжей, также как искусственная продукция ERC достаточна для индукции клеточного старения (Sinclair and Guarente, 1997). Итак, у дрожжей нестабильность рДНК явно коррелирует со старением, хотя ее роль в старении может быть и непрямой. Согласно одной из моделей высокое содержание ERC «вытитровывает» белки репарации и репликации ДНК из других участков генома и тем самым приводит к накоплению повреждений и/или уменьшению репликации генома.
Так как Sir2 является НАД-зависимой деацетилазой, а уровни НАД действуют как метаболический термостат, предположили, что влияние Sir2 на продолжительность жизни у дрожжей может быть связано с эффектами ограниченного питания, которое, как известно, замедляет старение у многих видов. Однако, хотя это предположение и подтверждается данными об увеличении активности Sir2 у дрожжей, дрозофилы и млекопитающих в условиях ограниченного питания, продолжительность жизни у дрожжей, выращиваемых в условиях пониженного уровня глюкозы (ограниченного питания), увеличивается механизмом, не зависящим и аддитивным по отношению к эффектам Sir2 (Kaeberlein et al., 2004). Следовательно, Sir2 и ограниченное питание повышают продолжительность жизни независимыми путями.
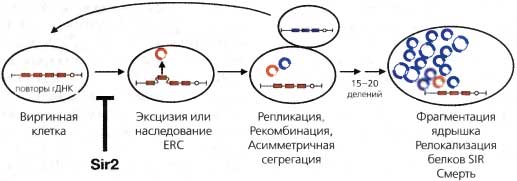
Рис. 4.11. Рекомбинация рДНК приводит к старению клеток у дрожжей
рДНК организована в массив из 140—200 прямых повторов 9.1-т.п.н. единицы {красный блок). Они кодируют 18S, 5.8S, 25S и 5S рРНК и содержат два Sir2-чувствительных элемента ниже гена 5S и внутри гена 18S. рДНК повторы имеют склонность вырезаться в стареющих клетках дрожжей и накапливаться в материнских клетках в виде кольцевых молекул (Kaeberlein et al., 1999). Это явление коррелирует с преждевременным старением. Sir2 препятствует ему, подавляя процессы неравной рекомбинации и вырезания
Накопление вырезанных колец рДНК не было обнаружено у каких-либо других видов. Однако для Caenorhabditis elegans и грызунов предполагалось, что уменьшение продолжителности жизни связано с другими видами нестабильности генома. Аналогично эффекту потери Sir2, приводящей к неравным обменам между сестринскими хроматидами, у дрожжей, возможно, что нарушения теломерного гетерохроматина у млекопитающих приводят к слиянию хромосом «конец-в-конец», тем самым нарушая способность клеток к делениям. И хотя пока неизвестно, влияет ли на эти механизмы белок Sir2 у млекопитающих, тем не менее роль геномной нестабильности как общего фактора старения весьма вероятна, как и специфическая роль нарушений в структуре гетерохроматина.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК