1.2. Генетическая идентификация группы Polycomb
У всех многоклеточных передне-задняя ось специфицируется посредством определенных паттернов экспрессии генов НОХ (рис. 12.2). У Drosophila во время эмбриогенеза активность произведенных матерью (т.е. унаследованных через ооцит) и произведенных в зиготе транскрипционных факторов генерирует специфическую комбинацию экспрессии НОХ, необходимую для каждого сегмента. Этот сегмент-специфичный профиль активности генов НОХ поддерживается на протяжении всего развития мухи, спустя долгое время после того как исчезли регуляторы ранней транскрипции. Когда функция генов НОХ была охарактеризована генетически, были изолированы многие trans-действующие регуляторы. Среди них Пам и Эд Льюс (Lewis, 1978) идентифицировали и генетически проанализировали Polycomb (Рс). Гетерозиготные мутантные по Рс самцы имели дополнительные половые гребешки на второй и третьей паре ног, тогда как самцы дикого типа несли половые гребешки только на первой паре ног (см. рисунок на титульной странице этой главы). Гомозиготные мутанты были летальны на стадии эмбриона, демонстрируя трансформацию всех кутикулярных сегментов в направлении самого заднего абдоминального сегмента (рис. 11.2в, г). Эти классические фенотипы PcG были интерпретированы как обусловленные эктопической экспрессией генов НОХ. Таким образом, Рс и другие гены со сходными фенотипами были определены как репрессоры активности генов НОХ. Впоследствии детальный анализ позволил обнаружить, что белки PcG требуются лишь для поддержания репрессии НОХ, а не для установления активности НОХ в ходе формирования паттерна, специфичного по отношению к положению Эта последняя задача выполняется транскрипционными факторами, закодированными в рано действующих генах сегментации. На основе их репрессирующего или активирующего влияния на экспрессию генов НОХ эти вновь идентифицированные trans-действующие регуляторы были разделены на два антагонистических класса, PcG HtrxG, соответственно (рис. 11.1) (Kennison, 1995).
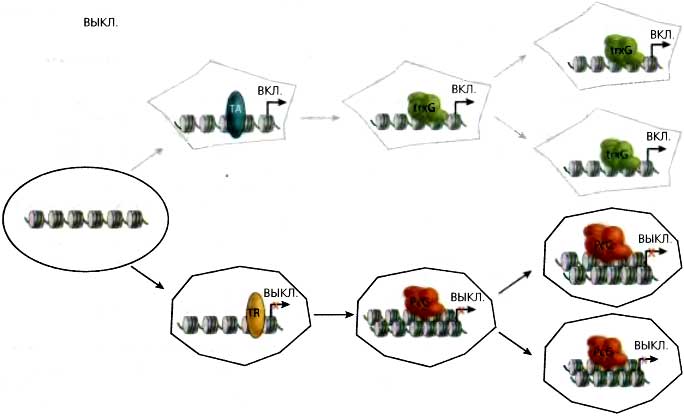
Рис. 11.1. Концепция клеточной памяти
Схема, иллюстрирующая участие комплексов PcG и trxG в детерминации активного и репрессированного состояний экспрессии генов и, тем самым, клеточной дифференцировки, которая поддерживается на протяжении многих клеточных делений: (ТА) транскрипционный активатор; (TR) транскрипционный репрессор
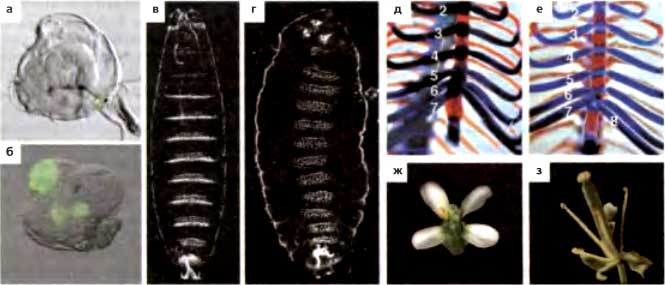
Рис. 11.2. Гомеотические трансформации у PcG-мутантов разных видов
(а-г) Drosophila melanogaster, (д, е) Mus musculus, (ж, з) Arabidopsis thaliana. (а, б) Ножные имагинальные диски, претерпевающие трансдетерминацию, на что указывает экспрессия крыло-специфического гена vestigial (маркированного GFP). (в. г) Кутикулы эмбриона дикого типа (в) и мутанта Su(z)12 (г). У мутантного эмбриона Su(z)12 все абдоминальные, торакальные и несколько головных сегментов (не все из них видны в этой фокальной плоскости) гомеотически трансформированы в копии восьмого абдоминального сегмента благодаря нарушенной экспрессии гена Abd-B в каждом сегменте, (д, е) Осевой скелет новорожденных мышей дикого типа (д) и RinglA-/- (е) Вид торакальных районов очищенных скелетов, показывающий кости (красный цвет) и хрящ (синий цвет). Мутант обнаруживает переднюю трансформацию восьмых торакальных позвонков, на что указывает наличие восьми (1—8) вертебростернальных ребер вместо семи (1—7), как у дикого типа, (ж, з) Цветок дикого типа (ж) и мутанта clf-2 (з). Цветок дикого типа демонстрирует нормальное расположение чашелистиков, лепестков, тычинок и плодолистиков. У цветка clf-2 лепестки отсутствуют или число их уменьшено. (а.б, с любезно предоставлено N. Lee and R. Раго; в, г, перепечатка, с любезного разрешения, из Birve et al., 2001 [©Company of Biologists Ltd.]; д, e, перепечатка, с любезного разрешения, из del Маг Lorente et al., 2000 [©Company of Biologists Ltd.]; з, любезно предоставлено J. Goodrich)
Молекулярное выделение генов PcG Drosophila сделало возможным изучение функции ортологов позвоночных у мышей, где они также оказались ключевыми регуляторами экспрессии генов НОХ (van der Lugt et al., 1994; Core et al., 1997). У млекопитающих мутации в генах PcG приводят к гомеотическим трансформациям позвонков (Рис. 11.2д, е). Кроме того, гены PcG играют ключевую роль в контроле клеточной пролиферации, поддержания стволовых клеток и рака (см. разделы 4.2 и 4.3 далее в этой главе). Замечательный консерватизм генов PcG у мух и млекопитающих облегчил биохимический анализ и привел к идентификации некоторых новых членов комплексов PcG, например белка RING 1 (Satijn and Otte, 1999). Направленная [targeted] мутация RING1 у мыши, например, привела к классической гомеотической трансформации фенотипа. Лишь впоследствии было обнаружено, что она соответствует гену Sex combs extra из группы PcG у Drosophila.
У двух других модельных организмов, а именно у червя Caenorhabditis elegans и цветкового растения Arabidopsis thaliana, молекулярная характеристика мутантов, изолированных в ходе разных генетических скринингов, выявила существование других ортологов белков PcG. У С. elegans члены группы PcG были идентифицированы в ходе скрининга на мутантов maternal effect sterile (mes), и было показано, что они участвуют в сайленсинге Х-хромосомы в зародышевой линии гермафродитов (Fong et al., 2002; глава 15).
У Arabidopsis гены PcG были идентифицированы в результате нескольких генетических скринингов при исследовании различных процессов развития (Hsieh et al/, 2003). Первый ген PcG у растений, CURLY LEAF (CLF), был идентифицирован как мутант с гомеотическими трансформациями органов цветка (рис. 11.2ж, з) (Goodrich et al., 1997). Мутации в генах класса FERTILIZATION-INDEPENDENT SEED (FIS) были обнаружены в ходе скрининга на мутанты, обнаруживающие материнский эффект—недоразвитие семян (Grossniklaus et al., 1998) или допускающие некоторое развитие семян в отсутствие оплодотворения (Luo et al., 1999; Ohad et al., 1999). Наконец, гены PcG были идентифицированы в результате скрининга на мутанты по срокам цветения, например мутанты, зацветающие сразу после прорастания (Yoshida et al., 2001) или с нарушенной реакцией на яровизацию — процесс, делающий растения компетентными к цветению после продолжительного воздействия холодом (Gendall et al., 2001).
Разнообразие процессов, регулируемых белками PcG, иллюстрирует значение поддержания репрессированного состояния ключевых регуляторов развития у различных организмов. С одной стороны, имеет место удивительный консерватизм некоторых биологических функций от растений до млекопитающих, например, регуляции таких ключевых регуляторов развития, как гомеотические гены, или участия в жесткой регуляции клеточной пролиферации. С другой стороны, комплексы PcG оказываются гибкими и динамичными молекулярными модулями, которые используются для контроля большой группы клеточных процессов и процессов развития.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК