4.1. Метилирование ДНК и мутации
Преимуществам метилирования ДНК как эпигенетической системы клеточной памяти противостоит недостаток, связанный с мутабильностью 5-метилцитозина. Цитозин (С) дезаминируется спонтанно, давая урацил (U), который затем неправильно спаривается с гуанином (Lindahl, 1974). Эта потенциальная мутация распознается урациловыми ДНК-гликозилазами, которые эффективно удаляют несоответствующее основание и инициируют репарацию, чтобы восстановить С на месте U. Однако, когда дезаминируется 5-метилцитозин, образуется тимин (Т). Результатом этого также оказывается неправильное спаривание, но тот факт, что Т, в отличие от U, является природным основанием ДНК, по-видимому, интерферирует с эффективным устранением повреждения. В результате мутантное тиминовое основание может сохраняться при репликации ДНК и передается дочерним клеткам как мутация типа транзиции С к Т. Мутации этого рода являются, по-видимому, одной из наиболее частых единичных причин генетического заболевания у людей, поскольку приблизительно одна треть всех точечных мутаций являются транзициями С к Т в последовательностях CpG (Cooper and Youssoufian, 1988). Эту нестабильность CpG в эволюционных масштабах демонстрирует далее 4—5-кратная недостача CpG в геноме млекопитающих (Bird, 1980). Единственным исключением оказываются островки CpG, в которых CpG не метилированы и поэтому стабильны.
Среди белков, связывающихся с метил-CpG, MBD4 остается пока уникальным в том отношении, что он обладает ферментативной активностью. Карбокситерминальный домен MBD4 является ДНК-гликозилазой, которая может in vitro избирательно удалять Т из неправильно спаренных Т—G (Hendrich et al., 1999). Эту активность можно было бы ожидать от системы репарации ДНК, которая исправляет дезаминирование 5-метилнитозина. В подтверждение этой гипотезы мыши, лишенные MBD4, обнаруживают повышенную мутабельность метилированных остатков цитозина в хромосомной репортерной последовательности (Millar et al., 2002). Кроме того, мыши, лишенные Mbd4, приобретают мутации типа транзиций С к Т в гене аденоматозного полипоза coli (АРС) и обладают повышенной частотой кишечных новообразований (табл. 18.2). Стоит отметить, что несмотря на существование специальной системы репарации, сайты метилирования цитозина сохраняются как «горячие точки» для мутаций.
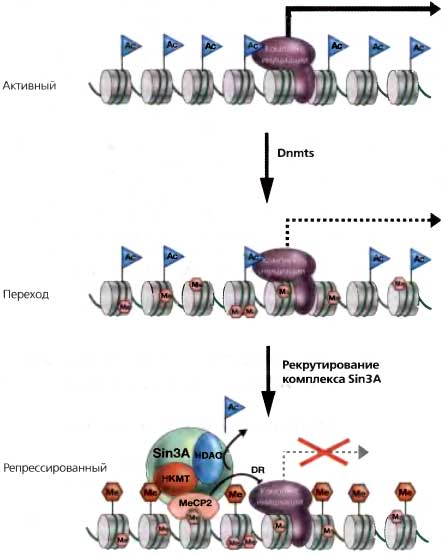
Рис. 18.6. Гипотетический переход между активным, неметилированным генным промотором и репрессированным промотором, «молчание» которого обусловлено метилированием ДНК, опосредованным МеСР2
Эта переходная фаза представляет собой промежуточный этап, во время которого транскрипция сайленсирована и происходит метилирование ДНК. Предполагается, что МеСР2 рекрутирует комплекс деацетилазы гистонов (HDAC) Sin3A и активность метилтрансферазы лизинов гистонов (НКМТ) к метилированным сайтам. Кроме того, имеются некоторые данные о том, что МеСР2 может прямо репрессировать транскрипцию путем контакта с комплексом инициации транскрипции (DR). Другие связывающиеся с метил-CpG белки взаимодействуют с разными корепрессорами, включающими активность НКМТ и (или) HDAC, и потенциально рекрутируют их
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК