3. Цитоплазматическая наследственность у инфузорий
Биология конъюгации P. aurelia чрезвычайно благоприятна для выявления эпигенетических явлений, связанных с влиянием цитоплазмы. В то время как реципрокное оплодотворение генерирует в двух конъюгантах генетически идентичные зиготические ядра, между ними почти не происходит обмена цитоплазмой, что позволяет эффективно различать действие ядерных генов от действия наиболее влиятельного фактора внешней среды — цитоплазмы материнской клетки (рис. 7.2). Каждое генетическое скрещивание поэтому эквивалентно изучению монозиготных близнецов, рожденных разными матерями. Исследования Соннеборна показали, что фенотипические различия между двумя родительскими клетками могут поддерживаться у потомков каждой из них даже несмотря на то, что эти потомки обладают идентичными генотипами. Позже оказалось, что в ряде случаев эти фенотипические различия детерминируются экстрануклеарными генами и сегодня не должны квалифицироваться как эпигенетические явления. Например, смертоносные свойства киллерных штаммов P. aurelia обусловлены обитающими в цитоплазме эндосимбиотическими бактериями рода Caedibacter; которые выделяют токсин, убивающий чувствительные штаммы (обзоры см. Preer et al., 1974; Pond et al., 1989). Другие случаи, такие как материнское наследование серотипа, которое требует взаимно исключающую экспрессию одного из нескольких паралогичных генов поверхностных антигенов, являются четкими случаями цитоплазматического влияния на активность генов.
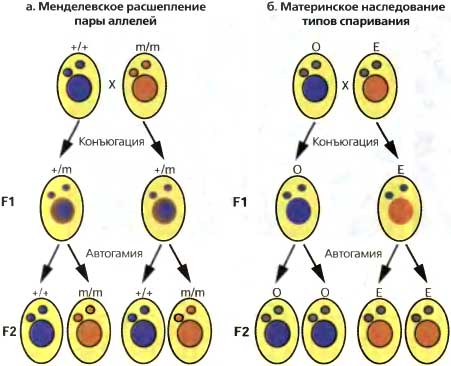
Рис. 7.2. Менделевская vs. цитоплазматическая наследственность
(а) Конъюгация и автогамия иллюстрируются скрещиванием между двумя клетками Paramecium, каждая из которых гомозиготна по разным аллелям, М или т. Конъюгация включает реципрокный обмен одним из двух идентичных гаметических ядер. В результате этого образуются клетки-эксконъюганты F1 с идентичными гетерозиготными генотипами. Автогамия (процесс самооплодотворения) генерирует полностью гомозиготный генотип всего в одном половом поколении, так что особи F2 имеют 50 %-ную вероятность стать М/М или m/m.
(б) Фенотипическое различие между клонами F1 выявляет цитоплазматически наследуемые признаки. У Paramecium тип спаривания (О или Е) необратимо детерминируется в ходе развития соматического макронуклеуса (большой кружок) из тотипотентного микронуклеуса зародышевой линии (маленький кружок); однако родительский макронуклеус направляет дифференцировку каждого эксконъюганта в направлении сохранения существующего типа спаривания
Как и серотипы, два комплементарных типа спаривания у P. tetraurelia, которые обозначаются как О и Е, демонстрируют цитоплазматический паттерн наследования. Признаки О и Е являются терминально дифференцированными фенотипами, которые детерминируются в ходе развития соматического макронуклеуса из тотипотентной зародышевой линии. После конъюгации вегетативный клон, происходящий от родителя О, почти всегда демонстрирует тип спаривания О, тогда как клон, происходящий от родителя Е, почти всегда относится к типу Е, несмотря на то, что оба этих эксконъюганта развиваются из идентичных зиготических геномов (рис. 7.26). Более того, когда между двумя конъюгируюшими клетками образуется большой цитоплазматический мостик, обусловливающий существенный обмен цитоплазмой, потомство обоих родителей обычно развивается как тип Е (Sonnebom, 1977). Таким образом, должен существовать некий цитоплазматический фактор, который направляет развитие типа Е.
Как описывается ниже, геном зародышевой линии во время развития макронуклеуса подвергается масштабным перестройкам ДНК, и хотя предполагаемый ген типа спаривания все еще не идентифицирован, было показано, что одна менделевская мутация, влияющая на детерминацию типа спаривания, нарушает эти перестройки генома в других локусах (Meyer and Keller, 1996). Если тип спаривания определяется регулируемыми геномными перестройками, этот цитоплазматический фактор, определяющий тип Е, должен обладать способностью направлять альтернативную перестройку гена типа спаривания и давать в результате макронуклеарную форму этого гена, специфицирующую тип Е. В ходе конъюгации эта форма должна также производить цитоплазматический фактор, определяющий тип Е и необходимый для дальнейшего наследования. Как описывается в разделе 10, паттерны альтернативных перестроек могут передаваться от материнских макронуклеусов к зиготическим макронуклеусам в результате транс-нуклеарного влияния, которое по-видимому опосредуется молекулами РНК, действующими зависящим от гомологии образом. Таким образом, цитоплазматический фактор, ответственный за неменделевское наследование, является, вероятно, молекулой РНК, контролирующей «мутацию» развития в гене типа спаривания.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК