1.7. «Молчащий» хроматин в центромерах необходим для опосредования когезии сестринских центромер и нормальной сегрегации хромосом
Как гетерохроматин центромерных внешних повторов и CENP-ACnp1-хроматин кинетохора влияют на общую функцию расхождения хромосом? Зависимый от Clr4 «молчащий» хроматин собирается на внешних повторах в центромерах, в родственном повторе в локусе типа спаривания и по соседству с теломерами. Эксперименты с «голыми» плазмидными ДНК-конструктами [naked DNA plasmid constructs] показали, что внешние повторы каким-то образом вносят свой вклад в сборку функциональной центромеры, наделяя эти Cen-плазмиды способностью сегрегировать на митотическом и мейотическом веретене. Но ни внешних повторов, ни центрального домена самих по себе недостаточно для сборки функциональной центромеры (Clarke and Baum, 1990; Takahashi et al., 1992; Baum et al., 1994; Ngan and Clarke, 1997; Pidoux and Allshire, 2004).
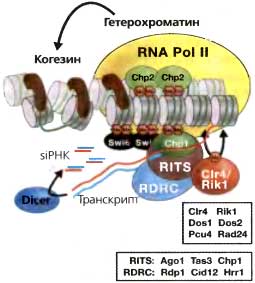
Puc. 6.6. Транскрипция центромерных повторов связывает RNAi, образование гетерохроматина и когезию
Транскрипция внешних повторов с помощью RNA pol II обеспечивает первичный субстрат для RNAi и зависимого от Dicer образования siRNA. Загрузка Ago1 в комплексе RITS (Ago1, Tas3, Chp1) молекулами siRNA делает возможным «нацеливание» гомологичного транскрипта. Действие RDRC (Rdpl, Cidl2 и Hrrl) делало бы возможной продукцию dsRNA, обеспечивая большее количество субстрата для Dcr1 для производства siRNA и, возможно, усиления сигнала. Взаимодействия между транскриптом, субъединицами RNA pol II и компонентами RNAi рекрутируют Clr4, который метилирует гистон H3 по лизину 9, делая возможным связывание хромодоменных белков, рекрутирование когезина и когезию сестринских центромер
Мутанты, вызывающие утрату сайленсинга во внешних повторах (т.е. дефектные по Clr4, компонентам RNAi или Swi6), повысили частоту утери хромосом в митозе и встречаемость отстающих хромосом на веретенах поздней анафазы (рис. 7а) (Allshire et al., 1995; Ekwall et al., 1996; Bernard et al., 2001; Nonaka et al., 2002; Hall et al., 2003; Volpe et al., 2003). Клетки без Swi6 дефектны по когезии в центромерах, но сохраняют когезию вдоль хромосомных плеч (Bernard et al., 2001; Nonaka et al., 2002). Формирование должным образом би-ориентированного веретена требует, чтобы сестринские кинетохоры прикреплялись к микротрубочковым фибрилам, отходящим от противоположных полюсов веретена. Силы, прилагаемые к би-ориентированным кинетохорам, требуют, чтобы сестринские кинетохоры прочно удерживались вместе (рис. 6.76). Swi6 необходим для рекрутирования когезина к хроматину внешних повторов и, тем самым, опосредует прочную физическую когезию между сестринскими центромерами (рис. 6.6). Таким образом, одна из функций «молчащего» хроматина в центромерах — быть посредником в когезии.
Когезин также прочно ассоциирован с теломерами и районом mat2-mat3 (Bernard et al., 2001; Nonaka et al., 2002). Кроме того, когезин также рекрутируется к «молчащему» хроматину, сформированному на гене ura+ в ответ на расположенный рядом эктопический центромерный повтор (рис. 6.5), подчеркивая связь между «молчащим» хроматином и когезией (Partridge et al., 2002). Таким образом, рекрутирование когезина, по-видимому, является общим свойством ассоциированного со Swi6 «молчащего» хроматина. Каким образом Swi6-хроматин осуществляет рекрутирование когезина — неизвестно, но Swi6 взаимодействует с субъединицей Psc3 когезина (Nonaka et al., 2002). Кроме того, Dfp1, регуляторная субъединица консервативной киназы Hskl (Cdc7), взаимодействует с Swi6 и требуется для рекрутирования когезина к центромерам (Bailis et al, 2003). Эта функциональная связь между гетерохроматином и когезией хроматид, по-видимому, сохраняется и у других организмов, поскольку уменьшение содержания компонента RNAi, Dicer, по-видимому, влияет на целостность гетерохроматина и когезию сестринских центромер в клетках позвоночных (Fukagawa et al., 2004).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК