1.2. Растения предоставляют дополнительные направления эпигенетических исследований
Наряду с общими для животных и растений элементами растения приобрели некие особенности, относящиеся к эпигенетическим феноменам. В то время как у животных оплодотворение осуществляется путем слияния двух гаплоидных клеток, которые возникли в результате непосредственно предшествующего мейоза, у растений существует гаплоидная (гаметофит) фаза роста между мейозом и оплотворением (рис. 9.1). Гаметофит представляет собой прорастающее пыльцевое зерно (мужская линия) и эмбриональный мешок (женская линия), каждый из которых имеет несколько гаплоидных ядер, которые образуются из мейотических продуктов вследствие двух или трех последовательных митозов, соответственно. На стадии гаметофита утрата генетической или эпигенетической информации не может компенсироваться информацией на гомологичных хромосомах или аллелях. Хотя исчерпывающие исследования в этой области знаний еще не проведены, тем не менее доказательств массивной утраты эпигенетических признаков во время гаметогенеза у растений, в отличие от животных, еще нет. Это может объяснить, почему у растений эпигенетические изменения часто передаются в результате мейоза.
Таблица 9.1. Сведения о геномных и эпигенетических компонентах у используемых эпигенетических модельных систем
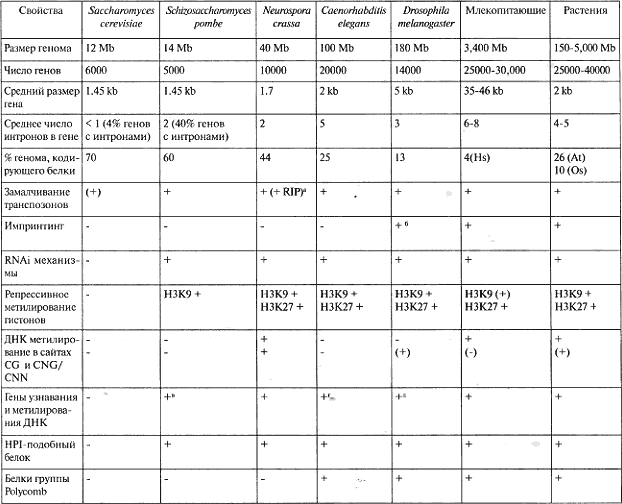
(At) Arabidopsis thaliana, (Cb) Caenorhabditis briggsae, (Ce) Caenorhabditis elegans, (Dm) Drosophila melanogaster, (Hs) Homo sapiens, (Os) Oryza saliva, (Pp) Pristionchus pacificus.
a Repeat-индуцированная точечная мутация, см. главу 6.
6 Хромосомная или геномная специфичность.
в Мутированная Dnmt2.
г Dnmt2 (Рр) и MBD-доменные белки (Се. СЬ, Рр). д Dnmt2 (Рр) и MBD-доменные белки.
Другая определенная отличительная черта растений — менее четко очерченная зародышевая линия и ее отделение от соматических клеток только на поздней стадии развития (рис. 9.1). У растений в точках роста на кончиках побегов и корней имеются апикальные меристемы, из которых образуются новые ткани и органы. Апикальная меристема побега дает начало цветкам, которые образуют гаметы для полового размножения, могут вырастать и дополнителные латеральные меристемы и образовывать цветы. У многих растений появились специализированные органы — ризомы, клубни или луковицы, которые имеют меристемы. Эти механизмы вегетативного размножения могут быть даже более общими и эффективными, чем размножение семенами. Зародыши могут образоваться не только в результате развития оплодотворенной яйцеклетки, но и из соматической ткани (соматический эмбриогенез). В тканевой культуре некоторые дифференцированные соматические клетки могут дедифференцироваться и перепрограммироваться в сторону альтернативной дифференцировки. Поэтому соматическое клонирование, пока еще мало эффективное у животных, широко распространено у многих видов растений, и с годами могут быть воспроизведены бесчисленные «зеленые Долли». Однако у клонированных казалось бы генетически униформных популяций растений набюдается выраженная фенотипическая изменчивость. Эта так называемая «сомаклональная вариабельность» имеет четкую эпигенетическую основу и потенциально может быть использована в селекции и адаптации растений (см. обзор Kaeppler et al., 2000)
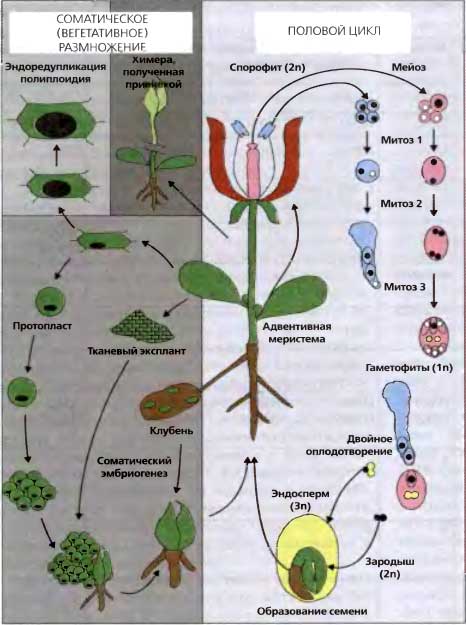
Рис. 9.1. Особенности жизненного цикла растения
Растения могут размножаться половым путем (образование гамет, оплодотворение и образование семян (правая часть рисунка) и бесполым (соматическим) путем (вегетативное размножение, де- и редифференцировка или эмбриогенез (левая часть рисунка). Тело высшего растения (корни, стебель, цветок) — диплоидный спорофит. При мейозе число хромосом уменьшается вдвое. Если у животных продукты мейоза образуют гаметы без дальнейшего деления и они сливаются с образованием диплоидного ядра, то у растений образуются гаплоидные мужской или женский гаметофиты в результате двух или трех митотических делений, соответственно. В конечном итоге пыльцевая трубка содержит одно вегетативное (белое) и два генеративных (черные) ядра. Два генеративных ядра оплодотворяют яйцеклетку (черная) и формируется центральная клетка с диплоидным ядром, произошедшим в результате слияния двух полярных ядер (желтые) Таким образом, в результате этого двойного оплодотворения и образуются диплоидный зародыш и триплоидный эндосперм, который снабжает развивающийся зародыш питанием. После прорастания семени зародыш дает начало новому спорофиту. Кроме того, большинство растений способны размножаться вегетативно благодаря активации покоящихся латеральных меристем, выросту специализированных корневых структур (клубни), амплификации в тканевых культурах или даже регенерации из индивидуальных соматических клеток после удаления клеточной стенки (протопласты). У растений часто наблюдается эндоредупликация, которая приводит к образованию полиплоидных клеток или тканей Химерные растения можно получить с помощью прививки. Поэтому генетическая и эпигенетическая информация у растений проходит гораздо меньше четко определенных путей развития зародышевой линии, чем у животных
Еще одна отличительная черта растений — существование плазмодесм (цитоплазматические мостики между клетками), которые проницаемы для небольших молекул, некоторых белков, РНК и вирусной геномной информации. Несмотря на большую взаимосвязанность, побеги растений можно срезать и привить на генетически отличные подвои (рис. 9.1). Это приведет к образованию химер, у которых вегетативные и генеративные ткани генетически различны. Поэтому хотя эпигенетические признаки и передаются по зародышевой линии, у растений они более изменчивы и обратимы, чем у животных.
Растения более толерантны к полиплоидии (умножение полного хромосомного набора), чем животные. У растений встречается множество диких и культурных полиплоидных видов (пшеница, хлопчатник, картофель, арахис, сахарный тростник, табак и другие). Полиплоидность дает им некоторые преимущества. При исследовании нуклеотидных последовательностей многих растительных геномов, включая маленький геном Arabidopsis thaliana, получены четкие доказательства дупликации исходного генома и генов. Даже диплоидные растения могут иметь полиплоидные клетки, которые возникают гораздо чаще, чем некоторые высокоспециализированные полиплоидные клетки у млекопитающих. Образование полиплоидов часто связано со значительными генетическими и эпигенетическими изменениями (см. обзор Adams and Wendel, 2005). Некоторые из этих изменений могут произойти на протяжении одного или нескольких поколений и они важны для быстрой адаптации и эволюции растений.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК