1.2. Способы определения пола
Генетические механизмы определения пола варьируют в значительной степени от одного организма к другому. Простейшая система включает один локус, который гомозиготен у одного пола (гомогаметныи пол) и гетерозиготен у другого (гетерогаметный пол) (рис. 17.1). Эта простая система развивалась различными путями, достигнув разной степени сложности у разных организмов. У некоторых из них возникли механизмы, подавляющие мейотическую рекомбинацию (кроссинговер) определяющих пол аллелей у гетерогаметного пола (рис. 17.1) — шаг, помогающий предотвратить возникновение смесей аллелей, ведущих к интерсексуальным состояниям. Во многих случаях неспособность к рекомбинации распространилась на часть или даже на всю хромосому, что сопровождалось утратой генетической информации. Эволюционное давление, обусловившее эту дегенерацию хромосомы, все еще остается непонятным, но конечный результат у многих видов заключается в том, что два пола обнаруживают различия не просто по аллелям в одном или нескольких локусов, но по целым хромосомам. Это изображено в виде простой диаграммы на рис. 17.1. У большинства видов, включая наш собственный, такую дегенерировавшую хромосому несут самцы, хотя бывают и исключения.
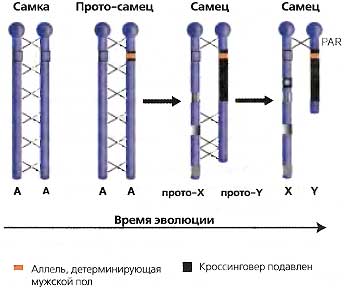
Рис. 17.1. Эволюция Y-хромосомы
На ранних этапах эволюции два пола могли различаться лишь по одному, аутосомному локусу, когда один пол (обозначаемый как протомужской) гетерозиготен по этому локусу, а другой пол (протоженский) гомозиготен. «Аллель, детерминирующая мужской пол», показана желтым цветом. Если для спаривания необходима одна особь каждого пола, тогда особи, гомозиготные по аллели, детерминирующей мужской пол, не могут возникать. На этой ранней стадии физиологические различия между полами будут еще слабыми, сравнимыми с различиями между двумя типами спаривания у дрожжей. Для предотвращения образования интерсексуальных состояний кроссинговер в локусе, определяющем мужской пол, и вокруг него будет подавлен (эти подавленные области показаны темным цветом) В пределах этого района будут накапливаться мутации, потому что подавление кроссинговера уменьшит их способность распространяться по популяции и, отсюда, давление отбора против них. Район дегенерации, в котором кроссинговер подавлен, будет постепенно расширяться («храповик Меллера»), пока эта хромосома не утратит большую часть своих активных, функциональных генов. Должен остаться небольшой, активный участок, гомологичный Х-хромосоме, для того, чтобы разрешить спаривание и кроссинговер при мейозе. Это псевдоаутосомный район (PAR, pseudoautosomal region). Аутосома, первоначально гомологичная будущей Y («А» на диаграмме), будет сама эволюировать в основном путем транслокаций с других аутосом. в конечном счете формируя отличающуюся Х-хромосому. Эта Х-хромосома, подобно другим хромосомам, представляет собой мозаику из фрагментов ДНК, помещенных на место в разные периоды на протяжении эволюции; некоторые из них являются древними, а некоторые относительно недавние. Это иллюстрируется по-разному расположенными пэтчами на прото-Х- и Х-хромосомах На Х-хромосоме человека более недавние участки обогащены генами, избежавшими инактивации X
Половая дифференцировка обычно запускается включением или выключением одного или небольшого числа генов в ходе развития. Продукты этих генов инициируют каскад регуляторных событий, опосредующих дальнейшее продвижение по тому или иному пути детерминации пола (см. детали для Caenorhabditis elegans в главе 15 и для Drosophila в главе 16). У человека существует белковый продукт гена SRYна Y-хромосоме, который запускает ранний зародыш по мужскому пути полового развития (обзор см.: McElrevey et al., 1995). Механизм такого типа не нуждается в крупных хромосомных различиях для того чтобы успешно работать; почему же тогда такие различия возникали так часто? Возможно, что они произошли как побочный продукт супрессии кроссинговера, необходимой для предотвращения интерсексуальных состояний (рис. 17.1). Математический анализ факторов, влияющих на распротстранение аллелей в популяциях, показывает, что супрессия кроссинговера неизбежно приведет к постепенному накоплению вредных мутаций (возможно даже, мутаций, вызывающих дальнейшее подавление кроссинговера). Это приведет к прогрессивной дегенерации одной из двух исходно гомологичных хромосом (рис
17.1). Этот процесс был назван «"храповиком" Меллера» в память генетика, впервые предложившего этот механизм. Для этого случая не существует отбора, он просто происходит как следствие начального принятия стратегии двух полов для воспроизведения (обсуждение см.: Charlesworth, 1996 и помещенные там ссылки). Какая бы эволюционная тенденция ни стояла за дегенерацией хромосомы, тот факт, что она произошла (и, возможно, еще продолжается), требует коэволюции механизмов, чтобы справиться с крупными хромосомными различиями между членами одного и того же вида. Для этого предназначены механизмы дозовой компенсации.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК