5. Эпигенетическая память
Известно, что в развитии позвоночных имеют место два рода эпигенетических модификаций генома. Это метилированный цитоцин во многих участках ДНК, где имеется CpG, и разнообразные модификации «хвостов» гистонов. Эти изменения приобретаются в ходе гаметогенеза и раннего развития и тесно связаны с активностью или неактивностью генов. Поэтому можно было ожидать, что эти эпигенетические модификации подвергнутся реверсии в результате пересадки ядра, а если нет, то могут помочь объяснить некоторые дефекты развития эмбрионов, полученных в результате трансплантации ядер.
У амфибий некоторое понимание деметилирования ДНК было достигнуто в экспериментах, где ядра соматических клеток млекопитающих были инъецированы в ооциты Xenopus (Simonsson and Gurdon, 2004). Промотор Oct-4 мышей метилирован в клетках взрослого тимуса, где Oct-4 не экспрессируется. Однако, когда ядра тимуса были инъецированы в ооциты, этот промоторный район, но не энхансерный район регуляторной части этого гена, был деметилирован. Этот результат показывает избирательный характер процесса деметилирования. Когда были инъецированы целые ядра, деметилирование ДНК, по-видимому, предшествовало индуцированной транскрипции Oct-4. Весьма вероятно, что активность ДНК-деметилазы является специальным свойством ооцитов (см. выше) и что деметилирование промоторной ДНК репрессированных в развитии генов может также быть важным и необходимым этапом, когда ядра соматических клеток репрограммируются в экспериментах с пересадкой ядер в яйцеклетки. Изменения в модификациях гистонов пока еще не были исследованы в экспериментах по пересадкам ядер у амфибий.
Эксперимент по пересадкам ядер у амфибий, поставленный по другой схеме, показал, что реверсия эпигенетического состояния соматических клеток никоим образом не наблюдается всегда. Ввиду невозможности получить ядерных трансплантантов у R. pipiens, даже от клеток-доноров с ранней стадии хвостовой почки (см выше), Бриггс и Кинг (Briggs and King, 1957) задались вопросом, не отражает ли морфология аномальных эмбрионов их происхождение; они описали предпочтительное выживание эндодермальных тканей у эмбрионов эндодермального ядерного происхождения и назвали это «эндодермальным синдромом». Указывалось, однако, что эндодерма дифференцируется позже, чем другие зародышевые листки, и что этим можно было бы объяснить ее лучшее выживание (Gurdon, 1963). Действительно, Симнет (Simnet, 1964) сообщил, что эмбрионы-трансплантаты нейрального происхождения также обнаруживали такое же предпочтительное выживание их эндодермы. Тот же вопрос недавно был снова исследован с использованием специфичных для клеточного типа маркерных генов. В этих опытах (Ng and Gurdon, 2005) ядра из клеток нейроэктодермы или эндодермы, уже экспрессирующих специфичные для клеточного типа маркеры Sox2 или эндодермин, соответственно, трансплантировали в денуклеированные яйцеклетки; получившиеся в результате эмбрионы-трансплантанты были разделены на нейроэктодермальную или эндодермальную части, и эти части были протестированы на те же маркеры, Sox2 или эндодермин (рис. 22.8 и табл. 22.4). Оказалось, что оба гена преимущественно экспрессировались в клетках «не своего» типа. Например, более половины эмбрионов нейроэктодермального происхождения обнаружили суперэкспрессию нейрального маркера Sox2 в своих эктодермальных клетках. У некоторых клонированных эмбрионов не было снижения уровня экспрессии гена Sox2 по сравнению с уровнем донорских клеток. Трансплантированные ядра, как и ядра нормальных эмбрионов, были совершенно неактивными в отношении транскрипции вплоть до стадии бластулы. Следовательно, как это ни удивительно, активное состояние генной транскрипции, установившееся в ходе клеточной дифференцировки, может поддерживаться в клонированных эмбрионах при полном отсутствии условий, которые индуцировали этот ген на протяжении более 12 митотических клеточных делений (от яйцеклетки до бластулы). Этот поразительный пример эпигенетической памяти наблюдается лишь у некоторых эмбрионов-трансплантатов, но полностью отсутствует у других, у которых экспрессия гена была успешно репрограммирована.
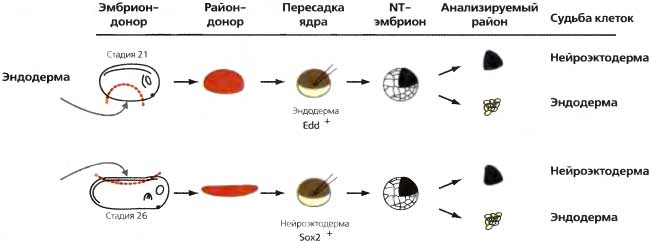
Рис. 22.8. Схема эксперимента по тестированию эпигенетической памяти об экспрессии генов, специфичной для данного клеточного типа
Донорские ядра берутся из клеток эндодермы (стадия 21) или нейроэктодермы (стадия 26). Получающиеся в результате пересадки ядер эмбрионы обычно формируют частичную бластулу, нормально дробившиеся части которой разделяют на будущие нейроэктодермальные или эндодермальные участки и анализируют по экспрессии генов. См. табл. 22.4 (перепечатано, с любезного разрешения, из Ng and Gurdon, 2005)
Табл. 22.4. Эпигенетическая память о клеточно-специфичной экспрессии генов
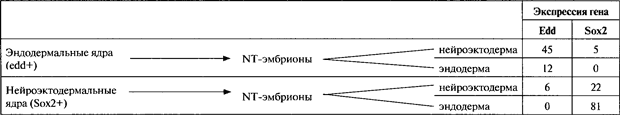
В процентах представлена доля эмбрионов, полученных путем трансплантации ядер (каждый тестировался методом RT-PCR индивидуально), экспрессирующих гены Edd и Sox2 в два и более раза интенсивней нормального (или фонового) уровня (данные из NG and Gurdon, 2005)
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК