2. Конъюгация: реципрокное оплодотворение обнаруживает неменделевскую наследственность
Инфузории являются гермафродитными линиями, способными к конъюгации — процессу спаривания, включающему перекрестное оплодотворение между двумя родительскими клетками. Зрелые клетки соответствующего клонального возраста, обычно после легкого голодания, становятся сексуально реактивными и спариваются с клетками совместимых типов спаривания, чтобы начать конъюгацию. Если совместимый партнер отсутствует, некоторые виды обычно претерпевают процесс самооплодотворения, называемый автогамией. В обоих случаях за этим следует ядерная реорганизация, начинающаяся с мейоза микронуклеусов. Последовательность ядерных событий сходна у всех видов (хотя и имеются некоторые вариации) и изображена на рис. 7.1 для видов Р aurelia и Т. thermophila. (см. Sonnebom 1975).
Постмейотическое развитие начинается с выбора единственного гаплоидного ядра в каждой клетке для передачи генома. Выбранное ядро претерпевает дополнительное деление, даюшее два генетически идентичных гаметических ядра. В случае конъюгации два партнера обмениваются одним из двух своих гаплоидных ядер, и поэтому последующая кариогамия (т.е. слияние двух гаплоидных ядер) формирует в каждом конъюганте генетически идентичные зиготические ядра (стадии 3—5 на рис. 7.1). При автогамии два гаметических ядра в одной клетке сливаются и образуют полностью гомозиготный диплоидный геном. В обоих случаях получающееся в результате диплоидное зиготическое ядро (стадия 5) делится еще два раза, и четыре продукта этих делений дифференцируются: два — в новые микронуклеусы, а два — в новые макронуклеусы (стадии 6 и 7). По завершении развития ядерного аппарата в новых вегетативных клонах сохраняются либо оба новых микронуклеуса, как это имеет место у видов P. aurelia, либо одно из двух ядер деградирует, как у Т. thermophila. У обоих видов два новых макронуклеуса при первом клеточном делении не делятся (стадия 9), а распределяются в две дочерние клетки; они начинают делиться только при втором вегетативном делении.
В то время как родительские микронуклеусы дают начало новым микро- и макронуклеусам следующего поколения, родительский макронуклеус утрачивается. У P. aurelia он фрагментируется примерно на 30 кусков, в которых репликация ДНК быстро подавляется, хотя транскрипция активно продолжается на протяжении периода дифференцировки новых макронуклеусов. Когда вегетативный рост возобновляется, эти фрагменты распределяются случайным образом в дочерние клетки, пока не остается ни одного (стадия 9). У Т. thermophila родительский макронуклеус не фрагментируется, но пикнотизируется и деградирует по апоптозоподобному механизму до первого вегетативного деления (Davis et al. 1992) (стадии 7 и 8).
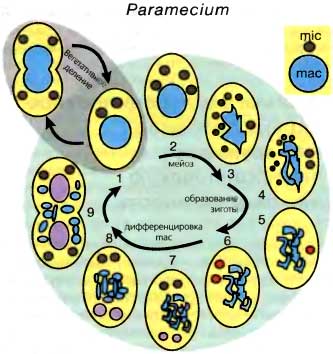
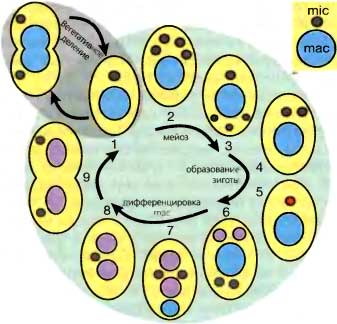
Рис. 7.1. Жизненные циклы Paramecium (вверху) и Tetrahymena (внизу)
(Стадия 1) Вегетативные клетки размножаются бинарным делением. Половое развитие (стадии 2—8) обычно начинается после конъюгации двух клеток или автогамии (только у Paramecium).
(Стадии 2—3) Мейоз микронуклеуса заканчивается выбором одного из гаплоидных продуктов на роль гаметического ядра и дегенерацией остальных ядер. У Parmecium родительский макронуклеус начинает формировать лопасти.
(Стадии 4—6) Формирование зиготы. Дополнительное деление выбранного ядра дает два генетически идентичных гаплоидных ядра. В ходе конъюгации одно из этих двух идентичных гаметических ядер обменивается на такое же ядро между двумя партнерами, и последующая кариогамия дает диплоидное зиготическое ядро (красный цвет). В ходе автогамии два идентичных гаметических ядра просто сливаются вместе. Два дополнительных постзиготических деления (6) дают недифференцированные микро- и макронуклеусы.
(Стадии 6—8) Ядерная дифференцировка. После второго постзиготического деления из получившихся в результате ядер два становятся новыми микронуклеусами, тогда как два других начинают дифференцироваться в новые макронуклеусы (розовый цвет). У Paramecium материнский макронуклеус фрагментируется. У Tetrahymena он пикнотизируется. У Tetrahymena же один из новых микронуклеусов дегенерирует.
(Стадия 9) Карионидное деление: Это первое вегетативное деление является особым, поскольку новые макронуклеусы распределяются в дочерние клетки без деления, тогда как микронуклеусы расходятся в клетки-потомки путем митоза. Наконец, фрагменты родительского макронуклеуса у Paramecium не делятся, но сохраняются до тех пор, пока не утрачиваются в результате случайного распределения при последующих делениях
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК