1.8. Эпигенетическое наследование функционального состояния центромеры
Интересное эпигенетическое явление было описано в отношении сборки функциональных центромер у дробянковых дрожжей на плазмидах, содержащих минимальные участки для центромерной функции. Хотя конструкты, сохраняющие лишь часть внешнего повтора и большую часть центрального домена, собирают функциональную центромеру неэффективно, но, как это ни удивительно, коль скоро это активное функциональное состояние центромеры установлено, оно может воспроизводиться в ряду многих митотических делений и даже в мейозе (Steiner and Clarke, 1994; Ngan and Clarke, 1997). Одно из объяснений этого сводится к тому, что внешние повторы обеспечивают среду, благоприятную для сборки кинетохора (Pidoux and Allshire, 2005), но, будучи собранным, CENP-ACnp1-хроматин (и, следовательно, кинетохор) воспроизводится в этом положении с помощью матричного механизма, который может быть сопряжен с репликацией (Takahashi et al., 2005). Возможно, что гетерохроматин каким-то образом индуцирует откладку CENP-ACnp1 в центральном домене или способствует ей (рис. 6.8) и что один только блок гетерохроматина не обеспечивает эффективную сборку кинетохора. Альтернативное объяснение заключается в том, что один внешний повтор не достаточен для рекрутирования достаточных количеств когезина, и это приводит к дефектной центромерной когезии и повышенным частотам утери хромосом. Такие центромерные конструкты после нескольких клеточных делений могут стохастически накапливать достаточные количества когезина, результатом чего оказывается увеличенная митотическая стабильность. Однажды достигнутое, это стабилизированное состояние должно каким-то образом дуплицироваться на дочерних молекулах, чтобы сделать возможным его воспроизведение в последующих делениях (рис. 6.8).
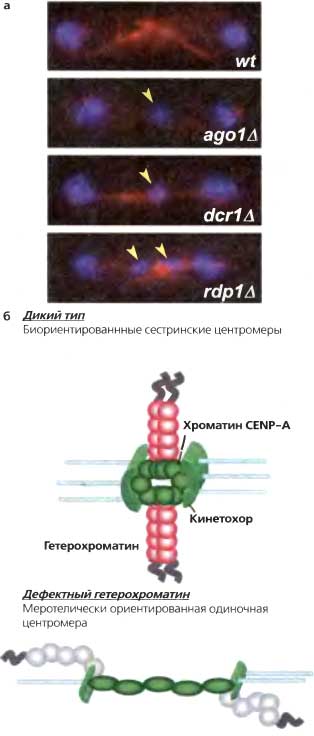
Рис. 6.7. Утрата гетерохроматина приводит к дефектному расхождению хромосом
(а) Клетки, лишенные RNAi или компонентов гетерохроматина, обнаруживают увеличенную частоту хромосомных потерь и отстающих хромосом на веретенах в поздней анафазе, (б) Отстающие хромосомы в клетках с дефектным гетерохроматином могут быть результатом дезорганизации кинетохоров, так что одна центромера может прикрепляться к микротрубочкам от противоположных полюсов. Такая меротелическая ориентация могла бы сохраняться при переходе в анафазу; разрыв прикрепления к одному или другому полюсу приводил бы к случайной сегерегации, результатом чего были бы случаи потери-приобретения хромосом
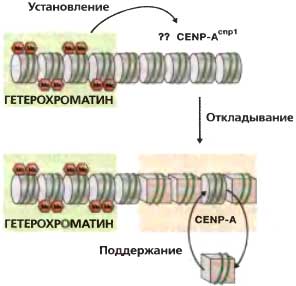
Рис. 6.8. Установление и поддержание хроматина CENP-A
ДНК центрального домена сама по себе не способна устанавливать функциональную центромеру; необходимы внешние повторы. Утрата гетерохроматина из установленных центромер не влияет на сборку CENP-ACnp1 или кинетохора на центральном домене. Это позволяет предполагать, что гетерохроматин каким-то образом указывает сайт CENP-ACnp1-хроматина и, тем самым, сборки кинетохора. Неизвестно, как CENP-ACnp1-хроматин откладывается в нуклеосомах или каким образом этот специализированный хроматин поддерживается в центральном домене
Как упоминалось выше, TSA также может устанавливать дефектное по центромере состояние, вызываемое утратой сайленсинга (Ekwall et al., 1997). С учетом связи между формированием «молчащего» хроматина и когезией в центромерах представляется вероятным, что индуцированное TSA гиперацетилирование блокирует эффективную реконструкцию «молчащего» хроматина посредством RNAi и что дефектная функция центромеры воспроизводится благодаря утрате сайленсинга и, таким образом, когезии сестринских центромер (дополнительные детали см. в главе 14).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК