3.4. Кластеры импринтированных генов содержат по меньшей мере одну ncRNAb
Из 12 известных кластеров импринтированных генов восемь ассоциированы с ncRNA (O’Neill, 2005). Раньше считалось, что ncRNAs, за исключением тех, которые участвуют в процессинге и трансляции РНК, т. е. таких как РНК сплайсинга, траспортные и рибосомные, являются раритетом в геноме млекопитающих. Теперь, благодаря доступности нуклеотидных последовательностей геномов мыши и человека, может быть выполнен анализ транскриптома, дающий перечень всех РНК-транскриптов в данной клеточной популяции. Уже показано, что большая часть транскриптома млекопитающих (не считая ncRNAs, связанные с процессингом и трансляцией) в основном, как это ни удивительно, состоит из ncRNAs, а не и PHК, кодирующих белки. Большой объем транскрипции ncRNAs в геноме млекопитающих и тот факт, что большое число ncRNAs перекрываются с известным кодирующим белок геном, показывают, что это не может рассматриваться как «транскрипционный шум», но представляет, вероятно, новую, до сих пор неизвестную систему регуляции генов (Mattick, 2005). Был создан Web-сайт, названный NON CODE, чтобы собирать данные о функциональных ncRNAs по всем организмам (http://noncode.bioinfo.org.cn/). Имеются несколько типов ncRNAs млекопитающих, которые, как было показано, обладают регуляторными функциями по отношению к генам, в том числе «короткие» ncRNAs от 21 до 30 п.о., участвующие в путях РНК-интерференции (глава 8), «межгенные» непроцессируемые транскрипты, регулирующие локальную активность хроматина (Haussecker and Proudfoot, 2005), и «длинные» процессируемые ncRNAs, такие как Xist, которая участвует в инактивации Х-хромосомы (глава 17).
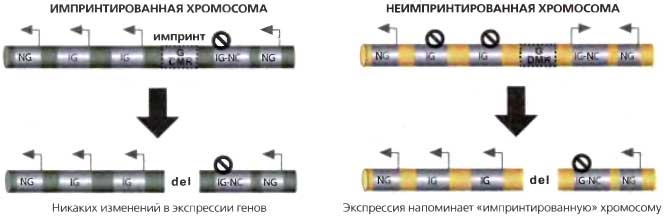
Рис. 19.6. Импринтированная экспрессия регулируется гаметическими DMR
На левой части рисунка показан эффект делегирования гаметического DMR из импринтированной хромосомы (зеленый цвет). Правая часть демонстрирует эффект делегирования гаметического DMR из неимпринтированной хромосомы (желтый цвет). Во многих импринтированных кластерах (например, lgf2r, Kcnq1 и Dlk1) экспериментальная делеция G-DMR влияет лишь на хромосому, несущую неимпринтированныйОМЯ. Результатом этого является утрата репрессии импринтированных генов иРНК, кодирующих белки (IG) и приобретение репрессии импринтированного гена ncRNA (IG-NC). Следует обратить внимание на то, что в некоторых импринтированных кластерах (Igf2 и Pws). которые здесь не изображены, метилированный G-DMR оказывается также необходимым для экспрессии некоторых из импринтированных иРНК в cis-конфигурации: (del) — делегированная ДНК, (G-DMR) — гаметический диффернциально ДНК-метилированный район, (NG) — неимпринтированный ген. (стрелка) — экспрессируемая аллель (черный запрещающий знак) — репрессированная аллель, (IMPRINT) — эпигенетическая модификация, ведущая к изменению экспрессии гена в cis-конфигурации
Какие типы ncRNAs связаны с кластерами импринтированных генов? Анализ ncRNAs, ассоциирующихся с шестью хорошо охарактеризованными импринтированными кластерами, перечисленными в табл. 19.2, все еще неполон, выявляя некоторые черты сходства, но и некоторые различия. Три импринтированные ncRNAs являются необычно длинными зрелыми РНК: Air имеет длину 108 т. о. (Lyle et al., 2000), Kcnq1ot1 — по крайней мере 60 т. о., но окончательный размер еще не определен (Mitsuya et al., 1999), a Ube3aas может превышать 1000 т. о. (Landers et al., 2004). Напротив, ncRNA Н19 имеет длину всего 2.3 т. о. (Brannan et al., 1990). ncRNA Gtl2 содержит множественные альтернативно сплайсированные транскрипты; однако была отмечена также межгенная транскрипция «вниз по течению», позволяя предполагать, что вероятны и более длинные транскрипционные единицы (Tierling et al., 2005). ncRNa Nespas крупнее, чем можно определить на РНК-блотах, и ее полный размер неизвестен (Wroe et al., 2000). Все эти импринтированные ncRNAs бедны интронами, имеют низкое отношение «интрон — экзон» или не сплайсированы как зрелые транскрипты. Ранее предполагалось, что все импринтированные гены бедны интронами; однако это может быть верно лишь для импринтированных ncRNAs (Hurst et al., 1996). Еще одной особенностью является то, что две импринтированные ncRNAs (Ube3aas и Gtl2-mpancKpunmbi «вниз по течению») действуют как хозяйские транскрипты для snoRNAs (малых ядрышковых РНК, которые направляют модификации к рРНК, snRNAs и, возможно, иРНК, действуя тем самым как посттранскрипционные регуляторы). snoRNAs не направляются к импринтированным генам иРНК в кластере, и вполне вероятно, что они не играют никакой роли в самом механизме импринтинга (Seitz et al., 2004). Аналогичным образом, miRNAs в кластере Dlk1 участвуют в посттранскрипционной репрессии одного из генов и РНК в кластере, но их роль в регулировании импринтированной экспрессии кластера еще не проверена (Davis et al., 2005).
Две особенности импринтированных ncRNAs показывают, что они могут играть роль в сайленсинге импринтированных генов иРНК (т. е. генов, кодирующих белки) в кластере. Первая заключается в том. что ncRNA обычно обнаруживает реципрокную специфичную в отношении родителя экспрессию по сравнению с импринтированными генами иРНК (табл. 19.2). Во-вторых, DMR, несущий гаметический импринт метилирования, который контролирует импринтированную экспрессию целого кластера, перекрывается с промотором ncRNA в трех случаях (Air, Kcnq1otl, Gnas Exon la). Это могло бы служить указанием на то, что эти импринты возникли для регуляции ncRNA в каждом импринтированном кластере В пользу такой интерпретации свидетельствуют эксперименты, в которых делетировали неметилированную последовательность, несущую гаметический DMR, вызывая тем самым утрату экспрессии ncRNA одновременно с приобретением экспрессии импринтированных генов иРНК (рис. 19.6), как было проверено в кластерах Igf2r, Kcnq1, Gnas, Pws и Dlk1 (Wutz et al., 1997; Bielinska et al., 2000; Fitzpatrick et al., 2002; Lin et al., 2003; Williamson et al., 2006). Кластер Igf2 представляет собой исключение, потому что оказывается, что неметилированный гаметический DMR не является непосредственным регулятором ncRNA Н19 (Thorvaldsen et al., 1998).
Эксперименты, в которых прямо тестируется роль самой ncRNA, были проведены сейчас для двух ncRNAs из кластеров импринтированных генов типа I (Air и Kcnq1otl) и одной ncRNA (Н19) из кластера типа II. Укорочение ncRNA>4/r длиной 108 т. о. до 3 т. о. показало, что сама ncRNA необходима для сайленсирования всех трех генов иРНК в кластере Igf2r, указывая тем самым на четкую регуляторную роль этой ncRNA (Sleutels et al., 2002). Кроме того, укорочение ncRNA Kcnq1otl, имеющей длину 60 т. о., до 1.5 т. о. также показало, что эта ncRNA прямо нужна для сайленсирования всех десяти генов иРНК в более крупном кластере Kcnq1 (Mancini-DiNardo, 2006). Напротив, точная делеция ncRNA Н19 и промотора не оказала никакого влияния на импринтинг в кластере lgf2 в тканях эндосперма, хотя в мезодермальной ткани наблюдалась некоторая потеря импринтинга (Schmidt et al., 1999). Таким образом, два из матерински-импринтированных кластеров типа I имеют общий механизм сайленсинга, зависящий от ncRNA, тогда как единственный отцовски-импринтированный кластер типа II, изученный до настоящего времени, использует другую модель, зависящую от инсулятора. Мы с нетерпением ждем результаты по другим импринтированным кластерам, чтобы посмотреть, указывает ли все это на существование только двух типов базовых механизмов импринтинга у млекопитающих — один для отцовски-импринтированных кластеров, а другой для матерински-импринтированных кластеров.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК