2.4. «Подавление» (quelling)
Вскоре после того, как для Neurospora были разработаны методы трансформации, исследователи в нескольких лабораториях заметили, что заметная доля (например, ~30 %) трансформантов Neurospora обнаруживают сайленсинг трансформирующей ДНК и, что еще более удивительно, сайленсинг нативных последовательностей, гомологичных последовательностям трансформирующей ДНК. Эта последняя форма сайленсинга в вегетативной фазе была названа лабораторией Макино (Macino) «подавлением» («quelling»). К настоящему времени эта лаборатория выполнила большую часть исследований этого феномена (Pickford et al., 2002). Подавление выглядит наиболее демонстративно с видимыми маркерами, такими как гены albino, которые кодируют ферменты, необходимые для биосинтеза каротиноидов (рис. 6.13). Полагают, что оно сопоставимо с «косупрессией» или _ посттранскрипционным сайленсингом генов (PTGS) у растений. Интересно, что гены, по-видимому, варьируют по своей чувствительности к подавлению. Для генов, которые чувствительны, подавление кажется самым обычным у трансформантов, несущих множественные копии трансформирующей ДНК, расположенные компактно. Ядра свободно плавают в гифах Neurospora, создавая ситуации «гетерокариоза», в которых генетически различающиеся ядра находятся в общей цитоплазме. Таким образом, легко показать, что «подавление» является «доминантным»; т.е. трансформированное ядро может сайленсировать гомологичные последовательности в соседних ядрах (Cogoni et al., 1996). Это предполагает наличие цитоплазматического фактора сайленсинга, возможно какой-то РНК. В согласии с этой возможностью идентификация генов, участвующих в «подавлении», показала, что «подавление» тесно связано с RNAi в других системах (Pickford et al., 2002). Говоря конкретно, qde-1, qde-2, qde-3, dcl-1 и dcl-2 кодируют, соответственно, RdRP, белок, подобный белку “Argonaute”, который, как полагают, участвует в направляемом малыми интерферирующими РНК (siRNA) расщеплении иРНК (Baumberger and Baulcombe 2005), RecQ-подобную презумптивную ДНК-геликазу (йву-3) и два «Dicer», которые предположительно участвуют в образовании siRNA (Catalanotto et al., 2004). Хотя критические детали пока отсутствуют, разрабатываемая модель сводится к тому, что в некоторых случаях трансформирующая ДНК генерирует «аберрантный» транскрипт, который каким-то образом включает машинерию RNAi, что ведет к деградации гомологичных иРНК. Хотя метилирование
ДНК часто связано с трансформирующей ДНК, ни метил-трансфераза ДНК, DIM-2, ни метилтрансфераза лизина 9 гистона H3, DIM-5, для подавления не нужны (Cogoni et al., 1996; Chicas et al., 2005).
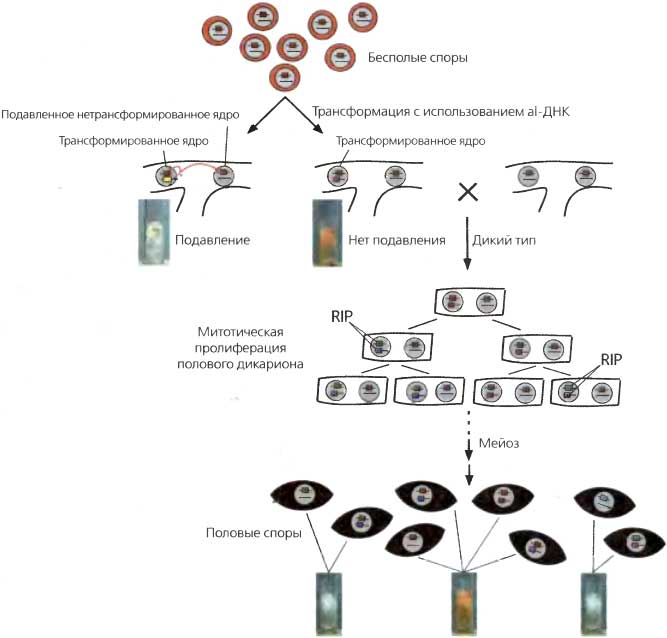
Рис. 6.13. Подавление (quelling)
Для простоты изображены лишь две из семи хромосомы (отрезки прямых линий в серых кругах, представляющих ядра). Нативный ген albino (al) показан темно-оранжевым прямоугольником на верхней хромосоме; другой (темно-оранжевый или желтый) прямоугольник представляет эктопические последовательности al, введенные путем трансформации. Поскольку трансформированные клетки часто являются многоядерными, трансформанты, как показано, часто бывают гетерокариотическими. Независимо от того, включает ли трансформирующая ДНК весь кодирующий район или нет, у некоторых трансформантов он сайленсирует («подавляет») нативный ген аl+ как в трансформированных, так и в нетрансформированных ядрах посредством неидентифицированной trans-действующей молекулы (красные линии, отходящие от трансформирующей ДНК, обозначенной желтым прямоугольником). Результатом этого оказывается слабопигментированная или альбинотическая (Аl-) ткань у некоторых трансформантов, как это показано на рисунке
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК