3.4. Регуляция случайной Х-инактивации — выбор
При некоторых обстоятельствах могут иметь место отклонения от случайной Х-инактивации. Это происходит либо в результате нарушений в первоначальном выборе
Х-хромосомы для инактивации («первичная неслучайная инактивация), либо в результате отбора против клеток, поддерживающих конкретную Х-хромосому в активном состоянии («вторичная» неслучайная Х-инактивация) (рис. 17.7). При первичной неслучайной Х-инактивации на выбор влияют вариации cis-действующих последовательностей или мутации, влияющие на вероятность того, что данная Х-хромосома будет выбрана у гетерозиготных животных как активная или неактивная X. Согласно предложенной Растаном модели блокирующего фактора, эти вариации могут проявлять свое влияние, изменяя вероятность связывания блокирующего фактора с данной аллелью.
Примером первичной неслучайной Х-инактивации является контролирующий Х-хромосому элемент (Хее, X controlling element) у мышей. Хее — классическим образом определенный локус, где разные аллели оказались влияющими на вероятность того, что несущая этот локус Х-хромосома будет активной X у гетерозигот XX (Cattanach and Isaacson, 1967). Опыты по генетическому картированию помещают Хее непосредственно «вниз по течению» от Xist (рис. 17.6) и, следовательно, в правильном положении для того, чтобы влиять на связывание блокирующего фактора, как определяется делецией 65 т. п.н. Лежащую в основе этого изменчивость последовательностей остается идентифицировать.
Второй элемент, который у мышей может влиять на выбор, — это антисмысловой регулятор Tsix (Lee and Lu, 1999). Полагают, что он опосредуется антисмысловой РНК Tsix, которая транскрибируется через локус Xist перед инициацией случайной Х-инактивации (рис. 17.6). Хромосома, несущая делецию промотора Tsix, в клетках XX, претерпевающих случайную Х-инактивацию, инактивируется предпочтительно. Более слабые отклонения наблюдаются у клеток с мутациями в энхансерных элементах (Xite-элементах), которые могут управлять уровнями экспрессии Tsix. Хотя промотор Tsix лежит в пределах района, определяющего предполагаемый сайт связывания блокирующего фактора, опыты с «прицельными» делециями не активируют Xist «по умолчанию» в ES-клетках XY. Это позволяет предположить, что эти локусы не синонимичны, но что делеция промотора Tsix оставляет интактными сайты связывания для блокирующего фактора.
Транскрипция Tsix сопровождается транскрипцией Xist, идущей на низком уровне, перед инициацией случайной Х-инактивации, позволяя предполагать, что влияние на выбор может быть опосредовано двунитевой РНК (т. е. гибридными нитями Tsix: Xist). В согласии с этим повышение уровня смысловой транскрипции через промоторы Xist находится в противоречии с репрессивным эффектом Tsix и дает в результате аллель с меньшей вероятностью стать активной X в гетерозиготной клетке XX (Nesterova et al., 2003). Подытоживая этот материал, можно сказать, что на рис. 17.6 показаны все различные Xic-элементы, о которых известно, что они влияют на инициацию случайной и импринтированной Х-инактивации, и которые часто описываются с точки зрения их способности влиять на функции счета и (или) выбора.
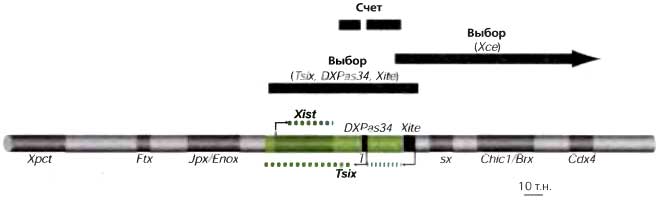
Рис. 17.6. Гены и регуляторные элементы в районе центра Х-инактивации
Ключевой район, регулирующим Х-инактивацию, изображен зеленым цветом. Фланкирующие гены показаны серым цветом Стрелки указывают на промоторы генов Xist (смысловой) и Tsix (антисмысловой). Протяженность соответствующих некодируюших РНК изображена прерывистыми зелеными линиями. Регуляторные элементы, контролирующие экспрессию Tsix, Xite и DXPas34, показаны черным. Районы и локусы, участвующие в выборе Х-хромосомы и счете Х-хромосом, показаны в верхней части рисунка
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК